Yacimientos líticos del Pleistoceno en la India. Petraglia 2007; Field et al, 2007. (1) Didwana Complex, (2) Budha Pushkar, (3) Hockra, (4) Luni Valley, (5) Hiran Valley, (6) Beas-Berach, (7) Bhimbetka, (8) Adamgarh, (9) Samnapur, (10) Patne, (11) Konkan Complex, (12) Upper Son, (13) Belan Valley, (14) Middle Son, (15) Dang-Deokhuri Complex, (16) Singbhum, (17) Ramayogi Agraharam, (18) Bora, (19) Kaladgi Basin, (20) Shorapur Doab, (21) Kurnool, (22) Attirampakkam, (23) Batadomba-Lena, (24) Fa Hien Cave, (25) Site 50, (26) Site 49, (27) Site 55.
Michael Petraglia y Nicole Boivin revisan las evidencias sobre los primeros poblamientos del sur de Asia.
Middle Son Valley, de tecnología achelense, ha sido datado en 140-130 ka (Petraglia et al, 2012) e indica la supervivencia de homínidos arcaicos, quizá Homo heidelbergensis (Athreya, 2007).
Las evidencias genéticas apuntan a una colonización de la región por el HAM hace 70-50 ka (linajes mitocondriales U2i, M2 y R5).
Los restos más antiguos de sapiens se hallaron en la Cueva Fa Hien de Sri Lanka (Deraniyagala, 1992), datados en 31 ka.
Hallazgos de tecnologías del Paleolítico Medio anteriores a la erupción del Toba, hace 74 ka, sugieren que el sapiens pudo haber llegado al sur de Asia con anterioridad, por rutas costeras y transcontinentales. Una hipótesis alternativa, es que sus autores fuesen neandertales (Mellars in Balter, 2010).
Se han identificado pocos yacimientos costeros. Es posible que la mayoría de los primeros asentamientos permanezcan actualmente inundados.
Paul Mellars et al (2013) apoyan una dispersión costera de los humanos modernos desde África oriental hasta el sur de Asia hace 60-50 ka, asociada con tecnologías análogas a la Howiesons Poort y tecnologías relacionadas, junto con una amplia gama de características "modernas" culturales y simbólicas. Las herramientas líticas datadas con anterioridad, corresponderían a neandertales o a poblaciones premodernas. Ajustan el reloj mitocondrial a las fechas por ellos propuestas.
Korisettar (2007) ha sugerido que una ruta transcontinental pudo haber resultado más favorable: las llanuras de la península disponen de fuentes abundantes de agua, materias primas líticas y recursos animales y botánicos. Los corredores de las migraciones humanas permanecen desconocidos.
Petraglia y su equipo, al contrario que Mellars, defienden que las primeras migraciones del HAM por el sur de Asia se corresponden con tecnologías del Paleolítico Medio.
16R Dune (Desierto Thar) es el yacimiento más antiguo del Paleolítico Medio, datado en 126-108 ka (Achyuthan et al, 2007). No ha sido descrito. Podría corresponder con Homo arcaico o con sapiens (Petraglia et al, 2010).
Se han identificado cientos de yacimientos en el subcontinente indio con métodos de reducción laminar, discoidal y levallois. Las herramientas son típicamente informales aunque están presentes hojas retocadas, buriles y puntas.
Los núcleos simples regulares y las técnicas discoidales están presentes ampliamente a lo largo de la India.
Los conjuntos de hoja son más comunes en el norte.
Los núcleos musteriense y levallois, en Pakistán, acompañados de una variedad de retocados, quizá correspondiendo con una ocupación neandertal.
Se han encontrado conjuntos del Paleolítico Medio datados en 78-38 ka en Jwalapuram, Jurreru River Valley (Petraglia et al, 2007, 2009). Los núcleos son similares a los de la MSA.
La capa pretoba de la localidad 3, está datada en 77 ka. Se hallaron 215 artefactos y un trozo de ocre rojo.
La capa posttoba, está datada en 74 ka, lo que demuestra que los seres humanos del Paleolítico Medio de India sobrevivieron a la erupción del Toba.
Lena Asryan, Andreu Ollé, Norah Moloney y Tania Rey presentan los artefactos líticos recuperados en las unidades V, III y II de la Cueva Azokh, Nagorno Karabaj, Cáucaso Menor, entre 2002-2009, datados en 293-100 Ka.
La entrada principal, Azokh 1, es una gran cueva que con dos secuencias geológicas (inferior y superior) y nueve unidades geo-arqueológicas de la que sólo las superiores (Unidades I a V) muestran un registro arqueológico significativo.
En 1968 se encontrón una mandíbula de homínido en la Unidad V, que fue datada en 250-400 ka (Guseinov, 1973, 1985). Fue clasificada como Palaeoanthropus azykhensis por Kasimova (1986, 2001) debido a que presentaba un mosaico de características arcaicas y derivadas. King et al (en prensa) sugieren que el fragmento es similar a los especímenes de Mauer, Arago y Atapuerca-SH, y lo incluyen tentativamente en Homo heidelbergensis.
Los artefactos encontrados en la cueva son principalmente productos finales dominados por herramientas en escama.
El conjunto de la Unidad V (68 piezas) se compone principalmente de escamas sin retocar con una presencia mínima de escamas y núcleos retocados. Datado en unos 300 ka.
Se hallaron cuatro lascas en la Unidad III.
El conjunto de la Unidad II (315 artefactos) incluye un componente importante de Levallois, aunque con un menor número de núcleos y lascas retocados. Se asocia claramente a Modo 3 o Musteriense. Datado en 184-100 ka.
En todas las unidades fueron explotadas diferentes materias primas locales y alóctonas. La obsidiana posiblemente proviene de fuentes distantes. La gama de materias primas es más variada en la Unidad II. Las materias primas de mejor calidad son las preferidas para las piezas retocadas.
Asociados a los restos líticos, se encontraron restos de fauna y hogueras.
Situación del Abrigo de la Quebrada y otros yacimientos valencianos
Aleix Eixea, Clodoaldo Roldán, Valentín Villaverde y João Zilhão han comparado las materias primas utilizadas en el Abrigo de la Quebrada con las de otros yacimientos valencianos del Paleolítico Medio mediante análisis macro y microscópicos.
Han identificado cuatro materias primas diferentes en los hallazgos del Abrigo de la Quebrada: pedernal, piedra caliza, cuarcita y cuarzo. El pedernal es la materia prima más abundante en todos los niveles, seguida en menor proporción por calizas y cuarcitas. El cuarzo, es de utilización esporádica.
En los alrededores del yacimiento, el pedernal tipo Domeño es el tipo de piedra dominante, con varios afloramientos cercanos. Ofrece buena calidad para la talla. Con respecto al pedernal alóctono, se han definido cuatro grupos, en base a la observación macroscópica, de fuentes desconocidas por el momento.
Los autores advierten que los autores son provisionales. De los análisis y comparaciones se desprende un nivel significativo de movilidad a través de los corredores del Turia, Plana de Utiel y Magro, que conectan el interior y la costa. Estos ríos convergen a la gran llanura de Valencia, lo que permite el movimiento hacia el Norte (Quebrada) y hacia el sur (Cova Negra, Cova Foradada, Cova del Gat, Cova de la Petxina, Cova Beneito, Abric del Pastor y El Salado). Para los autores, el patrón de distribución espacial observada refleja la circulación de los distintos grupos a través de unos territorios de subsistencia de tamaño más bien grande. Alternativamente, podría reflejar la circulación de materias primas a través de redes de intercambio de larga distancia.
Reconstrucción por ordenador de los cinco cráneos de Dmanisi. Imagen: Marcia Ponce de León y Christoph Zollikofer de la Universidad de Zúrich
Homo georgicus, Gabunia et al, 2002.
En 1991 Atje Justus halló en las excavaciones de la villa medieval de Dmanisi, Georgia, la mandíbula D 211 datada en 1,8-1,7 Ma. Con 16 piezas dentarias, únicamente le faltan las dos ramas ascendentes. Aspecto masivo y forma estrecha, fuerte reducción del espacio retromolar y ausencia de mentón.
Comparación de varios ejemplares de erectus y georgicus.
En 1999 se hallaron:
La calota completa D 2280. Conserva parte de la región basal del cráneo con las partes basal y nucal del occipital ligeramente dañadas, partes del ala grande del esfenoides y la mayoría de la fosa mandibular temporal.
Giorgy Nioradze halló el cráneo D 2282, calota y fragmento de maxilar. Deformado. Los maxilares retienen piezas dentales ligeramente desgastadas y alveolos de dientes que son visibles en radiografía. Posiblemente perteneció a una hembra joven.
Mandíbulas de Dmanisi. D 211, D 2600 y D 2735.
D 211 y D 2600 presentan polaridades molares opuestas
En 2000 se extrajo la mandíbula D 2600. Solo le falta el
extremo inferior de las ramas mandibulares. Muy grande, la mayor atribuida al
género Homo; por esta razón se ha
especulado con su pertenencia a otra especie diferente. Ramas mandibulares muy
altas. Incisivos de tamaño muy pequeño, especialmente los centrales. Caninos
grandes comparativamente. Premolares con dos raíces, lo cual es un carácter muy
arcaico. Molares grandes y masivos, de esmalte grueso. Polaridad opuesta a la
de D 211: tercer molar más grande que el segundo y este más que el primero.
Buena parte de estas características coinciden con las de habilis y rudolfensis,
pero el tamaño es mayor. Por ello, Leo Gabunia propuso la nueva especie Homo georgicus.
Para Skinner et al (2008) las diferencias entre D 2600 por una parte, y D 211 y D 2735 por la otra, son demasiado grandes para que se deban a dimorfismo sexual. José María Bermúdez de Castro ha apuntado que en este caso de hipotético dimorfismo sexual se daría la paradoja de que el individuo más grande, “el macho”, tendría una capacidad craneal mucho más pequeña que las supuestas hembras.
Para Ann Margvelashvili et al (2013), usando las poblaciones modernas de cazadores-recolectores como referencia, las variaciones en características como la forma de la arcada dental, la altura del corpus mandibular o la inclinación de la sínfisis, permanecen dentro del rango normal. El desgaste excesivo de los dientes conduce a la ruptura de los mecanismos normales de remodelación, lo que resulta en patologías, pérdida de dientes y pérdida de la función masticatoria y además en la extraordinaria altura de la mandíbula. Sin embargo, es poco probable que ello haya limitado la duración de la vida porque este efecto fue probablemente paliado por la preparación de alimentos blandos.
María Martinón-Torres (2013) señala la posibilidad de que esta mandíbula no sea patológica. En todo caso, las patologías habrían producido una recesión del borde superior de la mandíbula. Además resalta que del análisis del desgaste se deriva una alimentación herbívora, similar a la de chimpancés y gorilas. Esta dieta sustancialmente vegetal podría sugerir una adaptación a un nicho ecológico diferente para el individuo del cráneo 5 y los demás homínidos encontrados en el yacimiento. Además explicaría la ausencia de mecanismos compensatorios (como la migración dental mesial o mesial drift) habituales en Homo. En las mandíbulas D 211 y D 2375 se identifican estos cambios compensatorios, lo que evidencia un uso sustancialmente diferente de los dientes entre ambos conjuntos.
Laura Martín-Francés et al (2014) detectan un desgaste dental severo e inusual, acompañado de una extensa exposición de la raíz con migración axial, abscesos periapicales y fracturas en el esmalte. Además, hay evidencia de rotación dental post-eruptiva y artropatía temporomandibular. Proponen que el patrón de desgaste observado en este individuo, similar al registrado en chimpancés y gorilas y diferente al observado en el resto de Homo, se relaciona con una dieta con un alto consumo de alimentos fibrosos y abrasivos tales como frutas y vegetales. Las superficies oclusales redondeadas y las superficies labio-linguales muy pulidas de los dientes anteriores de D 2600 podrían ser principalmente la consecuencia de actividades pre o para-masticatorias, como la sujeción y la extracción. Este tipo de comida sería también el origen de la morfología oclusal en forma de copa de la dentición posterior en combinación con un relativamente ligero desgaste proximal. Las lesiones mostradas por D 2600 no han alterado significativamente la morfología de la mandíbula y no impiden una evaluación taxonómica apropiada.
Según Ann Margvelashvili et al (2016), las patologías son más prevalentes y el desgaste está más avanzado en las muestras de Dmanisi que en los actuales cazadores recolectores.
Para José María Bermúdez de Castro et al (2014), la combinación de rasgos arcaicos y derivados en D 2600 puede estar relacionada con la dieta. Las principales características derivadas de D 2600 con respecto a las otras mandíbulas, son las siguientes:
Presencia de un área retromolar subhorizontal y bien desarrollada.
TM 1517, Paranthropus robustus y D 2735 Homo georgicus
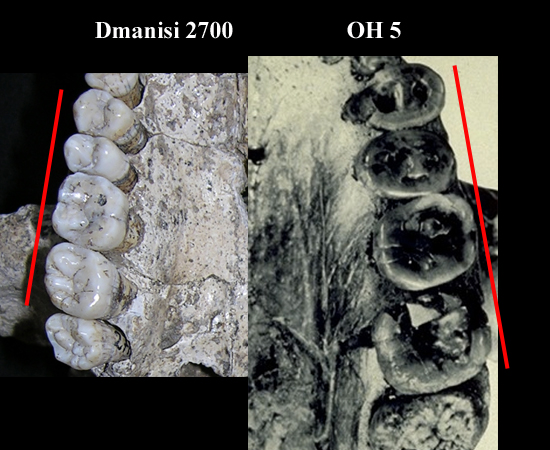
En 2001 apareció el cráneo D 2700 en extraordinario estado
de conservación y la mandíbula D 2735, completa, que encaja en el cráneo
anterior, que pertenecieron a un macho subadulto. Cráneo pequeño, dentro del
rango de habilis. Cara baja y
cóncava, de caninos prominentes y acusado prognatismo. Grandes caninos.
Incisivos grandes y en forma de paleta. Caninos grandes y prominentes.
Sts 5, Australopithecus africanus y D 3444, Homo georgicus
En 2002 Slava Ediberidze encontró el cráneo D 3444 de 650 cc, y la mandíbula D 3900, asociados a industria olduvaiense.
En 2003 se halló una mandíbula que recibió el nombre de Viejo de Dmanisi, ya que perteneció a un
anciano (40-50 años) que había perdido todos sus dientes hacía mucho tiempo,
por lo que debió de haber sido alimentado por otros humanos.
En 2005 se halló el Cráneo 5 (D 4500; David Lordkipanidze et al, 2013; Rightmire et al, 2017) que ha resultado corresponder al mismo individuo que la mandíbula D 2600. Esta combinación mezcla de forma inaudita un cerebro pequeño (540 cc) con una cara muy prognática y exhibe afinidades morfológicas con los Homo tempranos de África. Este individuo sería un varón que medía entre 1,40 y 1,60 m y pesaba alrededor de 50 kg. Cuando murió debía tener unos 30 años.
En 2011 se encontró un hueso de dedo del pie.
Además se han hallado un fémur, un peroné, varias tibias,
vértebras, costillas y diversos huesos de la mano y el pie. Las proporciones
entre los brazos y las piernas son claramente modernas. El pie muestra un arco
bien desarrollado, con el dedo gordo en posición paralela. El peso máximo sería
de 50 kg y la altura máxima 1,50 m. Las palmas de las manos estaban orientadas
hacia delante, lo que se interpreta como una adaptación a la vida arbórea.
Vistas de los cráneos de Dmanisi. Escala 5 cm. De arriba a abajo:
Escama del temporal baja, con el borde superior recto.
Punta de la mastoides envuelta y aplanada posteriormente.
Ángulo petrotimpánico bajo (140º-150º).
Pirámide petrosa lisa, densa en apariencia.
Foramen lacerum restringido.
Silla nasal prominente.
Posibles sinapomorfías con Homo erectus asiáticos.
Aplanamiento parasagital (depresión) de las superficies parietales.
Crestas paramastoidea y occipitomastoidea presentes.
Posibles autapomorfías.
Doble quilla sagital.
Placa timpánica relativamente delicada.
Proceso supratubario de Weindenreich ausente.
M3 es de tamaño reducido.
SK 847 (arriba), D4500 (medio) y AL 666-1 (abajo). Las flechas señalan similitudes. Escala 5 cm. David Lordkipanidze et al, 2013
Interpretaciones filogenéticas
La presencia de estos restos en Georgia nos indica que la
primera salida homínida de África, corrió a cargo de una especie de cerebro
pequeño, próxima a H. habilis.
Para Emiliano Aguirre (2008) los seres de Dmanisi
pertenecen a un grupo que salió de África en el momento de la cladogénesis del ergaster.
Parte distal del corpus de D 2735 (izquierda) y D 2600 (derecha)
María Martinón-Torres (2006), tras estudiar exhaustivamente la evolución del aparato dental de los homínidos, es partidaria de una nueva especie Homo georgicus antecesora de ergaster y erectus.
En 2008 su equipo analizó al detalle la dentición de los homínidos de Dmanisi, destacando el aspecto marcadamente primitivo de algunos de los dientes georgianos, en el caso de los caninos, semejantes incluso a especímenes de H. habilis o incluso del género Australopithecus. Los dientes de D 2600 tienen las coronas muy gastadas y su longitud, número y grado de bifurcación y molarización es marcadamente robusto y primitivo y, en su conjunto, solo presente en algunos especímenes de Homo rudolfensis, Australopithecus o incluso Paranthropus. En las otras dos mandíbulas, en D 211 y D 2375, a pesar de los rasgos habilinos, se dan unas características sorprendentemente derivadas (o “modernas”) para una población de esta antigüedad. Los dientes de D 211 y D 2375 presentan una serie molar decreciente (el primer molar es mayor que el segundo que es a su vez mayor que el tercero) y otras características relacionadas con un tamaño dental reducido que son inusuales para una población de esa antigüedad. Sugieren la posibilidad de que en el hipodigma de Dmanisi hubiera representadas al menos dos paleodemes o poblaciones. De la misma opinión son Jeffrey H. Schwartz, Ian Tattersall y Zhang Chi (2014).
Para Pontzer et al (2011), los molares de los seres de Dmanisi son similares a los del ergaster. La dentadura no estaba especializada lo que indica que georgicus explotaba una amplia gama de alimentos.
David Lordkipanidze et al (2013) suponen que los cinco cráneos hallados en Dmanisi se depositaron en un intervalo de unos 20 ka y pertenecen a un mismo demo muy variable. Ello daría pie a reconocer un rango muy amplio de variación morfológica en los primeros Homo y por consiguiente reunirlos en un único linaje evolutivo (habilis, rudolfensis, ergaster y erectus) (Rightmare et al, 2017).
D4500 y D2600 comparten características con todas las especies Homo tempranas, lo que sugiere un único linaje evolutivo muy variable.
Esta interpretación está siendo muy discutida. Parece difícil que no se haya producido especiación en demos muy reducidas, aisladas durante centenares de miles de años y explotando entornos muy diferentes. José María Bermúdez de Castro (2013) apunta que D 4500+D 2600 representa el cráneo más pequeño de la muestra al lado del aparato masticador más robusto, lo que induce a pensar que pertenece a otra especie diferente.
Si la mandíbula D 2600 no es patológica se hace difícil de explicar su relación con el resto de la muestra.
Ubicación estratigráfica de los hallazgos de Dmanisi
Calvo-Rathert (2008) sugieren que la acumulación de los homínidos de Dmanisi podría cubrir un periodo de hasta cientos de miles de años. La mandíbula D2 600 proviene de una capa con polaridad normal (es decir, pertenece al subcrón Olduvai y por lo tanto tiene más de 1,78 Ma) y algunos de los demás fósiles se han encontrado en capas con polaridad magnética reversa (post-Olduvai y por lo tanto más jóvenes que 1,78 Ma) (María Martinón-Torres, 2013).
Los fósiles se hallaron principalmente en dos áreas diferentes. Bloque 1 (D 211, D 2280, D 2282) y Bloque 2 (D 2735, D 2700, D 4500 y D 2600) separados de 10 a 20 m. Varios fósiles (D 2280, D 2282, D 211, D 2700 y D 2735) se atribuyeron a la unidad estratigráfica A, de polaridad normal (subcrón Olduvai), pero más tarde Ferring et al comunicaron que todos los fósiles humanos habían sido recuperados de los estratos inversamente polarizados B1x-B1z.
Un metatarsiano de adulto se halló en B1z
D 2700 y D 2735, se asignan a B1x.
D 2600, D 4500, D 3900 y D 3444 se asignan a la unidad B1y.
D 2280, D 2282 y D 211 no han sido atribuidos de forma específica a ninguna unidad estratigráfica.
Varios investigadores, como Lee Berger o María Martinón-Torres, han destacado la necesidad de comparar los ejemplares de Daminisi con Paranthropus y Australopithecus.
José María Bermúdez de Castro et al (2014) apoyan la existencia de dos paleodemos diferentes en Dmanisi: D 211 y D 2735 estarían viuculados a Homo habilis o Homo ergaster, mientras que las características en mosaico de D 2600 no se encuentran en ningún otro homínido y justifican la independencia del taxón Homo georgicus.
Herramientas líticas
Se han encontrado 2.241 objetos líticos, de más de 2 Ma. Los homínidos de Dmanisi explotaron fundamentalmente las rocas locales, sobre todo las procedentes de lechos fluviales. Cantos rodados y bloques angulosos de basalto, andesita y toba, formaron parte de los principales soportes utilizados. Gran parte de ellos, no fueron modificados.
El 82,7% corresponde a bases positivas o lascas de los que solo un 4,5% sobre el total corresponde a objetos retocados de pequeño formato. Un 33,2% de las lascas son corticales. Los núcleos representan un 4,3% del total y los choppers y chopping-tools un 10,4%.
Los análisis petrográficos demuestran que hubo traslado de ciertos materiales hasta el propio yacimiento, lo cual indica una cierta complejidad en el proceso de adquisisicón de las materias primas. La talla está completamente condicionada por la geometría natural de los soportes, a menudo ortogonal con ángulos de percusión muy altos, si bien se presenta una cierta diversidad de sistemas de talla. Los núcleos, lascas y debrises presentes, demuestran además que todo el proceso de reducción se elaboró en el mismo sitio. Numerosos núcleos unifaciales sugieren que no se trataba de una talla “demasiado elaborada”. También se documenta talla centrípeta en algunos núcleos sobre lasca (flake-cores). Sin bifaces. Predominio absoluto de lascas frente a herramientas. El sitio es comparable a Bed I de Olduvai o a Atapuerca TD-6.
El clima de Dmanisi, por entonces, debió ser moderadamente árido, con altas montañas surcadas por profundos valles y cuadalosos ríos, con paisajes abiertos de estepa cálida en las llanuras circundantes surcadas por bosques-galería en los valles. Por consiguiente, los homínidos de Dmanisi tuvieron a su disposición una considerable diversidad de hábitats.
Javier Baena ha identificado dos distintos conjuntos culturales.
Un hueso de venado hallado junto al cráneo 5 conservaba una escama lítica incrustada; marcas de herramientas en algunos otros huesos de animales, sugieren el consumo humano. Pero, por otra parte, los carnívoros parecen ser los agentes primarios de las acumulaciones fósiles.
Mandíbulas de Dmanisi. La flecha de arriba a la izquierda señala una lesión producida por el uso continuado de palillo.
La salida de África.
La misma crisis climática que condujo a la aparición del ergaster obligó al habilis y a otros seres parecidos a migrar siguiendo las masas de
bosque. Algunas poblaciones llegaron probablemente al Centro de África, donde
todavía hoy se mantienen los bosques tropicales. El bosque tropical es poco
propicio para la conservación de fósiles. Otras poblaciones se dirigieron hacia
el Norte, siguiendo las riberas del Nilo y el Mar Rojo. En el Cáucaso
encontraron un hábitat idóneo.
En 2015, Wei Qi comunicó en nota de prensa una datación de 1,95-1,77 Ma para el yacimiento de Heitugou, cuenca del Nihewan, Yangyuan, China, que sería por tanto anterior a Dmanisi.
Detalle de la parte izquierda de D 2600, donde se precibe una raiz molarizada bifurcada en P3
Extracción del cráneo 5.
Adam van Arsdale: Homo georgicus.
Bibliografía:
Gabunia, Leo, et al. "Dmanisi and dispersal." Evolutionary Anthropology: Issues, News, and Reviews 10.5 (2001): 158-170.
Vekua, Abesalom, et al. "A new skull of early Homo from Dmanisi, Georgia."Science 297.5578 (2002): 85-89.
Rightmire, G. Philip, David Lordkipanidze, and Abesalom Vekua. "Anatomical descriptions, comparative studies and evolutionary significance of the hominin skulls from Dmanisi, Republic of Georgia." Journal of Human Evolution 50.2 (2006): 115-141.
Lordkipanidze, David, et al. "Postcranial evidence from early Homo from Dmanisi, Georgia."Nature 449.7160 (2007): 305-310.
Molares rectangulares largos y estrechos en D 2735; redondeados en D 211.
Metacónido en P1 inferior de D 2735, ausente en D 211.
En P1, Entocónidos en D 2600, ausentes en D 211 y D 2735. D 2600 con tres largas raíces robustas, mientras que D 211 y D 2735 con una sola raíz, con ranuras por encima de una punta bífida.
Caninos inferiores orientados oblicuamente, ovoides en sección transversal en D 2600; bucolingualmente comprimidos en D 2735.
El M 3 de D 2600 difiere del de D 211 en tamaño y forma y en un gran, más mesialmente extenso entocónido y un gran hipocónido, más bucalmente expandido. Las raíces del M3 de D 2735 son menos abocinadas que las del M1, mientras que las de D 2600 están excepcionalmente bien separadas y las raíces de M2 y M3 de D 211 son individuales.
En el cráneo (D 211 está asociada con D 2282, y D 2600 con D 4500.
Supraorbitales arqueados no sobresalientes en D 2735; finos, rectos, anteriormente sobresalientes en D 2282; altos, en forma de barra, superiormente expandidos, con surco posttoral en D 4500.
Estas diferencias hacen que el conjunto aparezca visual y morfológicamente incoherente. Los autores proponen una identidad propia para D 2600, diferente a la de los otros ejemplares.
Comparación de varios ejemplares de erectus y georgicus.
José María Bermúdez de Castro et al (2014), apoyan también la posibilidad de dos paleodemos diferentes en Dmanisi: Según ellos, D 211 y D 2735 estarían viuculadas a Homo habilis u Homo ergaster, mientras que las características en mosaico de D 2600 no se encuentran en ningún otro homínido y justifican la independencia del taxón Homo georgicus.
Esta hipótesis ha sido rechazada anteriormente sobre la base de que todos los hallazgos corresponden a un mismo estrato y proceso tafonómico. D 2600 se ajusta al cráneo D 4500. David Lordkipanidze et al,(2013) consideran que la variabilidad del hipodigma de Dmanisi se puede alojar en un único linaje evolutivo (H. erectus georgicus ergaster) y esto implica que H. habilis, H. rudolfensis, H. ergaster y H. erectus son variantes regionales de un mismo linaje.
El papel de los carnívoros en TD6-2 parece haber tenido un impacto en el conjunto original después de que los homínidos hubiesen extraído una gran cantidad de nutrientes de los cadáveres.
Los restos de homínidos canibalizados no muestran marcas de dientes de carnívoros y tienen una mayor presencia de huesos de difícil conservación en comparación con los restos de ungulados. Esto muestra la capacidad de aquellos homínidos para controlar los recursos y mantener alejados a los competidores.
Los resultados sugieren que TD6-2 representa una sucesión de asentamientos con características diferentes.
La diversidad genética de los neandertales era menor que la de los seres humanos de hoy en día y el patrón de variación y acumulación de alelos deletéreos sugiere que las poblaciones de neandertales eran pequeñas y aisladas. El apareamiento entre familiares era mucho más común que entre los sapiens.
Los genes implicados en la morfología esquelética han cambiado más de lo esperado en el linaje evolutivo Neandertal (sobre todo, la curvatura lumbar, hiperlordosis; genes CUL7, GLB1, COL2A1, HSPG2, VPS13B y NEB) mientras que los genes implicados en la pigmentación y el comportamiento (agresividad e hiperactividad) han cambiado más en el linaje humano moderno.
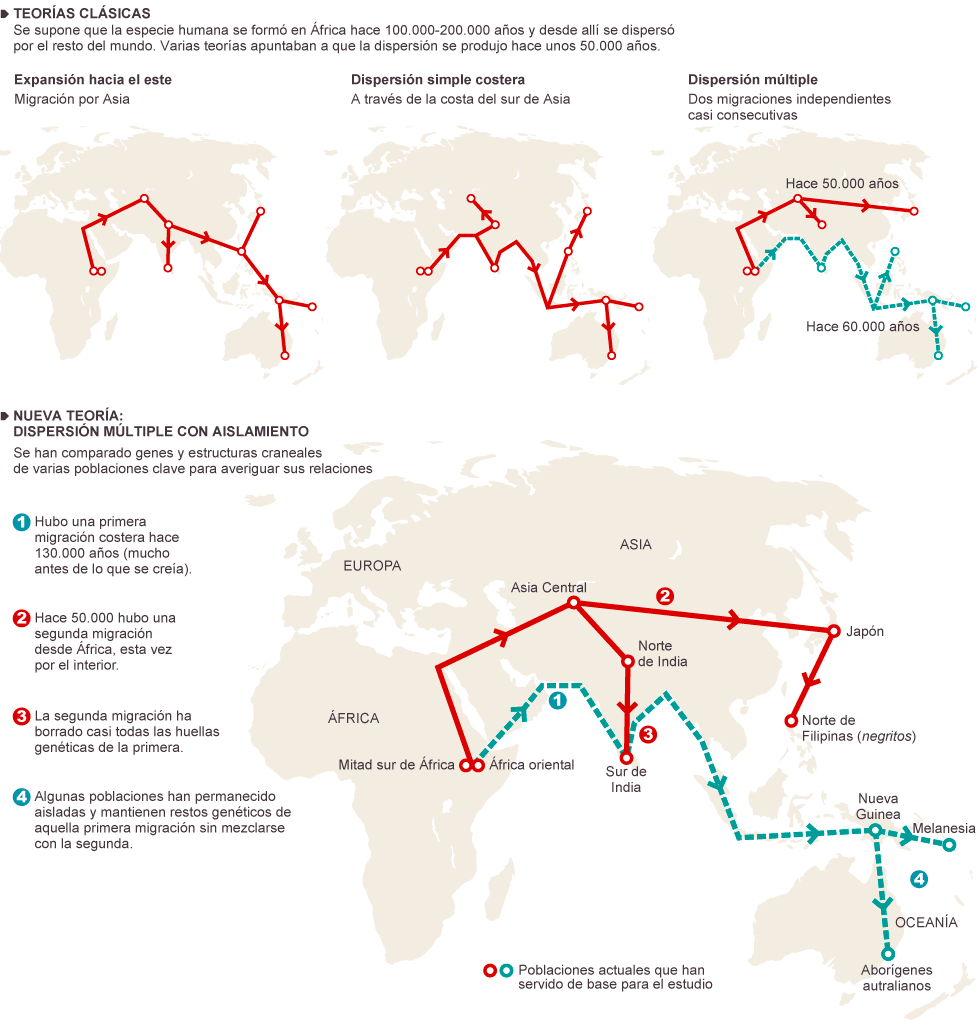
Una dispersión inicial por Asia a través de una ruta por el sur, por el estrecho de Bab el-Mandeb, hace ~130 ka, mucho antes de lo que hasta ahora se pensaba, de la que solo se conservan huellas genéticas y morfológicas en los aborígenes australianos y melanésicos.
Una dispersión posterior por el Corredor Levantino hacia el norte de Eurasia hace ~ 50 ka, seguiría preferentemente la ruta europea y un nuevo camino hacia Asia, pasando por el norte de la cordillera del Himalaya.
Karin Isler y Carel P. Van Schaik revisan las hipótesis que explican el crecimiento del cerebro humano a lo largo de la evolución en base a beneficios cognitivos. Las autoras sugieren que una perspectiva energética ayuda a complementar estos enfoques.
Alex Mackay, Brian A. Stewart y Brian M. Chase han analizado el registro arqueológico de África Meridional utilizando diferentes componentes de los sistemas tecnológicos líticos para rastrear la transmisión de información cultural y el grado de interacción de la población dentro y entre las diferentes regiones climáticas del Pleistoceno Tardío. Los datos sugieren un complejo conjunto de fusiones y relaciones entre poblaciones fragmentadas, con la máxima interacción durante MIS 4 y MIS 2 y máxima fragmentación durante MIS 5 y MIS 3. Los momentos de fusión de grupos se correlacionan con aumentos en la frecuencia de los ornamentos y otras formas de expresión simbólica, que llevan a los autores a sugerir que la interacción de la población fue un importante motor en su emergencia.
Los resultados demuestran el efecto de una homogeneización causada por la expansión austronesia de hace aproximadamente 3,5 ka. Los haplogrupos papúes autóctonos apoyan la hipótesis de una larga historia con algunos linajes con una profundidad temporal de 60 ka.
Distribución de DMRs a lo largo de las regiones genómicas y los sistemas corporales
La metilación nació evolutivamente como un sistema para inactivar transposones, antiguos genomas de virus que han perdido su capacidad infectiva pero conservan la de moverse de un sitio a otro por el genoma y sirve además para inactivar otras regiones del ADN. La ontogenia comienza con la mayoría de los genes abiertos, y el desarrollo del embrión implica la desactivación progresiva de unos genes u otros en cada zona del cuerpo.
David Gokhman, Eitan Lavi, Kay Prüfer, Mario F. Fraga, José A. Riancho, Janet Kelso, Svante Pääbo, Eran Meshorer y Liran Carmel han reconstruido los mapas completos de metilación del ADN de las muestras de hueso de neandertales y denisovanos, mediante el aprovechamiento de los procesos de degradación natural de citosinas metiladas y no metiladas. Comparando estos antiguos mapas de metilación a los de los seres humanos de hoy en día, se identificaron aproximadamente 2.000 regiones diferencialmente metiladas (DMR).
En particular, encontraron cambios sustanciales de metilación en HoxD9 y HoxD10 que pueden explicar las diferencias anatómicas entre los humanos arcaicos y actuales, en particular, las superficies de unión más amplias y relativamente más grandes y ejes de huesos curvos y miembros más cortos de los neandertales.
Otros cambios se relacionan con el desarrollo del cerebro, el sistema inmunológico y el cardiovascular. Muchos de los genes cuya actividad es exclusiva de los humanos modernos están vinculados a enfermedades como el alzheimer y la esquizofrenia.
El sistema digestivo se mantuvo relativamente sin cambios.
Además, hallaron una alta asociación de DMR y enfermedades.
Este estudio nos permite conocer el paisaje epigenético de nuestros parientes evolutivos más cercanos y abre una ventana para explorar los epigenomas de especies extintas.
En 23 artefactos, el desgaste de los bordes pudo atribuirse a la transformación de materias específicas.
En 7 casos (30%), procesamiento de tejido animal, corroborando la evidencia zooarqueológica de carnicería en el sitio.
En 16 (70%), tratamiento de tejidos vegetales, incluyendo madera, órganos de almacenamiento subterráneos, y tallos de hierba o juncos. En algún caso, podrían representar la producción y/o mantenimiento de herramientas de madera.
Michael Haslam ha revisado la información disponible sobre el uso de herramientas por los pánidos y los homínidos.
El ancestro común más reciente de seres humanos y pánidos, utilizaba herramientas, pero probablemente no hizo uso de las herramientas de piedra, y este rasgo surgió por primera vez entre los homínidos en África Oriental durante el Plioceno.
El uso de herramientas de piedra parece haber sido inventado independientemente por los chimpancés de África Occidental (P. t. verus) en la región de la moderna Liberia-Costa de Marfil-Guinea, posiblemente hace 200-150 ka. Por tanto, el ancestro común más reciente de bonobos y chimpancés tampoco utilizaba herramientas líticas.
Algunos IBD se comparten con el genoma ancestral reconstruido de los humanos y otros primates.
Otros IBD se comparten con neandertales o denisovanos. Muestran tramos más cortos en comparación con los segmentos no compartidos.
Muchos IBD compartidos con los denisovanos se encuentran exclusivamente en los asiáticos y son más largos que los segmentos compartidos con los denisovanos por otras poblaciones continentales. Por lo tanto, se confirma una introgresión denisovana en los antepasados de los asiáticos después de su migración fuera de África.
Los asíáticos comparten más IBD con los neandertales, pero los europeos comparten también con estos un porcentaje considerable de segmentos. Los IBD compartidos por neandertales y asiáticos o europeos actuales, son más largos que los compartidos por los africanos. Esto sugiere un flujo genético de los neandertales a los antepasados de los asiáticos y los europeos después de que salieron de África.
Muchos IBD compartidos por los africanos y los denisovanos o los neandertales son exclusivos en aquellas poblaciones y son sorprendentemente cortos. Por lo tanto, se supone que son muy viejos. Esto puede indicar que estos segmentos se derivan de ancestros de los HAM, neandertales y denisovanos que han sobrevivido en los africanos.
La evidencia de FwJj20 sugiere que los recursos acuáticos pueden haber desempeñado un papel importante en las dietas de principios del Pleistoceno y estos recursos pueden haber sido pasados por alto en las interpretaciones anteriores del comportamiento homínido. Los autores enfatizan el valor de la grasa en la dieta durante la estación seca FwJj20 presenta la evidencia más temprana de la explotación repetida de peces y tortugas por los homínidos.
Penny Spikins, Gail Hitchens, Andy Needham y Acebo Rutherford han estudiado las evidencias biológicas, culturales y sociales relativas a la infancia de los neandertales. Según sus conclusiones:
La niñez neandertal era sutilmente diferente a la de los humanos modernos, más enfocada a las relaciones sociales dentro del grupo. Los grupos neandertales estaban formados por menos individuos y vivían relativamente aislados.
Los niños neandertales, como se refleja en unos enterramientos más elaborados que los de los adultos, pueden haber desempeñado un papel particularmente importante en la sociedad, especialmente en el dominio de la expresión simbólica.
Los niños neandertales utilizaban el juego para desarrollar habilidades.
Es posible que los niños neandertales fuesen educados en la producción de herramientas. En un yacimiento de Francia y otro de Bélgica, se hallaron herramientas líticas hábilmente elaboradas junto a otras que estaban desconchadas torpemente.
Surco interproximal y estrías visibles en la superficie distal del P3 superior derecho de Yiyuan Sh.y.003. Las áreas señaladas en B se magnificaron en C y D.
(A: imagen óptica; B, C, y D: Imágenes de mocroscopia electrónica de barrido).
Chengkai Sun, Song Xing, Laura Martín-Francés, Christopher Bae, Liqun Liu, Guangbiao Wei y Wu Liu han analizado con microscopio binocular y microscopia electrónica de barrido las superficies interproximales de siete dientes fósiles descubiertos entre 1981-1982 en Yiyuan (Shandong, este de China), asignados a erectus.
Las coronas y raíces muestran diferentes grados de desgaste, desde estrías finas a ranuras interproximales profundas. La ubicación, morfología y dimensión de estos surcos o estrías son similares a las causadas por la utilización habitual de palillos de dientes.
Las características morfológicas que exhiben las marcas de los dientes de Yiyuan, las más antiguas de Asia, coinciden con las marcas de palillo ya identificadas en Homo habilis,Homo heidelbergensis y Homo neanderthalensis.
Análisis de componentes principales (bgPCA) de las mediciones angulares de la orientación del laberinto de los canales semicirculares. Los humanos (•) y los bonobos (▴) se solapan y están distantes de los chimpancés (+). Las elipses representan la dispersión que abarca aproximadamente el 67% de los sujetos.
Marwan El Khoury, José Braga, Jean Dumoncel, Javotte Nancy, Remi Esclassan y Frederic Vaysse han comparado la orientación del laberinto en los canales semicirculares mediante 260 tomografías computarizadas de humanos actuales (70 hembras y 67 varones), 61 bonobos (Pan paniscus) y chimpancés (8 Pan troglodytes verusy 54 Pan troglodytes schweinfurthi), todas las especies representadas por individuos subadultos y adultos.
Los humanos y los bonobos mostraron más similitudes entre sí que con los chimpancés. Este hallazgo no encaja con la bien establecida monofilia de chimpancés y bonobos.
Podría ser el resultado de una evolución convergente (homoplasias) en respuesta a presiones selectivas similares posiblemente asociadas con adaptaciones posturales.
Hay evidencia de una bipedación diferente en bonobos y chimpancés. Pontzer et al (2014), sugieren que la bipedación pudo haber sido relativamente costosa para los bonobos, en comparación con los chimpancés. Este costo puede reflejar una diferencia en la mecánica de la marcha.
La explicación más parsimoniosa es que bonobos y humanos retienen la morfología primitiva, mientras que la de los chimpancés es derivada.
El ángulo entre el plano medio sagital del cráneo y la orientación lateral de los canales semicirculares no muestra diferencias significativas entre las tres especies. Esto puede reflejar una condición primitiva compartida.
Por último, estos resultados podrían ser una consecuencia de la diversidad genética y la clasificación incompleta del linaje (A. Hobolth et al, 2010).
Sólo el ángulo entre el plano medio sagital del cráneo y la orientación anterior de los canales semicirculares, discrimina claramente a los humanos de las dos especies de Pan. Esto puede ser debido al hecho de que los bonobos y los chimpancés muestran características anatómicas que favorecen la versatilidad locomotora.
Otro hallazgo del estudio es la notable simetría de los canales semicirculares, con posibles aplicaciones en tafonomía.