lunes, 30 de septiembre de 2013
Nuevos enfoques para los estudios genéticos de las poblaciones antiguas
Pontus Skoglund esboza nuevos enfoques para la recuperación, análisis e interpretación de datos genómicos a gran escala de poblaciones antiguas, incluyendo soluciones a los problemas asociados con la cobertura limitada del genoma, la contaminación moderna, las diferencias temporales entre las muestras y los daños post-mortem en el ADN.
sábado, 28 de septiembre de 2013
Morfología de Homo ergaster y Homo erectus
 |
| Morfología del cráneo del erectus. Henke y Hardt. 2011. |
Rightmire (1990) recuerda que el diagnóstico formal de una especie debe basarse en sus rasgos apomórficos y no en los plesiomórficos.
- Por tanto, la bipedia no nos sirve como característica definitoria, como había propuesto Dubois, pues hoy sabemos que este rasgo es primitivo.
El grado erectus conserva otras plesiomorfias:
- Proyección hacia fuera de la cara (prognatismo facial), aunque
menor que en el habilis.
- Estrechamiento marcado detrás de los ojos.
- Frente huidiza.
- Molares relativamente grandes, aunque menos que los
australopitecinos. M1>M2>M3. Este último empiza a faltar.
- Caninos algo proyectados.
- Ausencia de barbilla.
Los verdaderos rasgos
distintivos, se relacionan con el cráneo (Stringer, 1984; Wood, 1984)
- Capacidades craneales 700-1.250 cc,
superiores al habilis. Esta es la característica morfológica más relevante, ya que duplica el volumen de alrededor de 500 cc, o poco más, propio de los Homo habilis y está muy por encima también de Kenyanthropus rudolfensis. El descubrimiento de restos postcraneales tan completos como el niño de Nariokotome, ha llevado a matizar el sentido evolutivo de ese aumento cerebral, entendiendo que en parte no sería sino una consecuencia lógica del aumento del tamaño de todo el cuerpo. Una expansión alométrica así rebaja un tanto el alcance de las hipótesis acerca del crecimiento relativo del cerebro en el Pleistoceno Medio, pero no hace sino confirmar el proceso evolutivo del linaje humano en esa época: hubo presiones selectivas a las que los Hominini respondieron aumentando de tamaño. Por otra parte existen evidencias muy ilustrativas de que las técnicas de construcción de útiles de piedra fueron ganando en complejidad, cosa que lleva a entender que no fueron sólo los cráneos, sino también las propias capacidades cognitivas, quienes sufrieron un incremento notable. Entre los ejemplares tempranos de África (850 cc) y los tardíos de Asia (1.150 cc) se da un incremento de un 25%.Mientras la caja craneal y el encéfalo se expanden, el rostro y el sistema masticador se reducen.
- Forma del cráneo característica
- La anchura máxima de la caja craneal está siempre situada hacia su base.
- Bóvedas aplastadas (platicefalia).
- Torus angular en la parte posterior e inferior del occipital. Cresta occipital. Hueso occipital muy angulado.
- Paredes craneales gruesas como consecuencia del comienzo de la inclusión de oquedades minúsculas, que permiten ganar espesor sin aumentar de peso (díploe, tejido esponjoso, neumatización) (Copes y Kimbel, 2015).
- Presenta algún carácter derivado ausente en el sapiens como una protuberancia horizontal en la parte posterior del cráneo (torus occipital), que le proporciona una apariencia alargada visto de lado y una protuberancia supraorbitaria bastante grande. Estas adaptaciones tienen que ver con un mayor papel de los incisivos en la masticación.
- La cara cuenta con un arco supraciliar grueso y continuo, un verdadero torus supraorbital aplanado por su parte superior. Rostro ancho y aplastado. Prognatismo menor que en los habilis. Las líneas temporales sobre las que se insertan los músculos masticadores temporales, están situadas en posición menos elevada que en los australopitecos.
- En los primeros erectus, las mandíbulas son relativamente más ligeras, con el sulco extramolar más estrecho que el de los Homo tempranos. La sínfisis es más delgada anteroposteriormente, el pozo geniogloso es relativamente bajo, y el planum postincisivo, aunque orientado oblicuamente, no es tan pronunciado como en otros Homo tempranos.
- Desarrollo de incisivos y caninos en detrimento de premolares y molares, lo que indica un régimen más omnívoro. Los dientes no difieren en tamaño absoluto a los de los Homo tempranos, pero sí en algunas proporciones y por ello son relativamente fáciles de diferenciar:
- El erectus muestra zonas oclusales más pequeñas que las de otros Homo tempranos.
- Raíces más pequeñas que las de otros Homo tempranos, pero más complejas que las de los HAM.
- Coronas relativamente más grandes. Especialmente en los molares, suelen ser bucolingualmente más estrechas en comparación con la longitud y menos bulbosas que en los Homo tempranos.
- Reducción de tamaño a lo largo de la fila molar con M3 reducido o similar en tamaño a M2.
- El erectus muestra una superficie articular de los huesos largos agrandada, huesos corticales gruesos, sobre todo en las extremidades inferiores, y un fémur anteroposteriormente aplanado, tróclea distal del fémur profunda, dobles meniscos anexos a la parte proximal de la tibia, pelvis reorientada menos amplia pero con mayor capacidad, marcado pilar ilíaco y torsión medial de la tuberosidad isquial.
- Las clavículas fósiles de erectus se encuentran dentro del rango normal de variación humana moderna. Esto apoya una reconstrucción del hombro de H. erectus similar al del HAM, con una cavidad glenoidea en posición lateral, lo que sugiere que la capacidad de lanzamiento se remonta a hace casi 2 Ma (Neil T. Roach y Brian G. Richmond, 2014; Neil T. Roach el al, 2013).
- Los cuerpos vertebrales lumbares son pequeños en relación con el peso corporal, a diferencia de los HAM. Los restos sugieren una posición primitiva del diafragma. El número de vértebras lumbares parece ser de cinco.
- Otras características torácicas son derivadas. El tórax es ancho en la parte superior y estrecho en la inferior.
- Los restos óseos sugieren un intestino relativamente pequeño, lo que tiene implicaciones para la calidad de la dieta y los posibles cambios de alimentación.
- La anchura bi-ilíaca, es similar a la de los australopitecos (Simpson et al, 2008; Ruff, 2010).
- Carol Ward V. et al (2013) han estudiado KNM-WT 51260, un tercer metacarpiano hallado en Kaito (oeste del Lago Turkana), datado en 1,4 Ma. El hueso se asemeja al de un humano moderno en proporciones generales y morfología. Es el tercero más largo de los Hominini conocidos anteriores a los neandertales y humanos modernos tempranos. En particular, KNM-WT 51260 muestra una apófisis estiloides bien desarrollada, una característica distintivas de la mano moderna y neandertal Neandertal, no presente en los primeros Hominini. Esta proyección ayuda a estabilizar la muñeca cuando la mano está agarrando pequeños objetos entre el pulgar y los dedos.
KNM-WT 51260 muestra que una morfolgía y función de la mano modernas estaban presentes dentro del contexto de la tecnología achelense y sugiere que las actuales características carpometacarpianas evolucionaron temprano, seleccionadas para un mejor manejo de las herramientas. - Hasta hace poco, se pensaba que el dimorfismo sexual era menor en erectus que en Australopithecus, pero la evidencia actual sugiere que pudo haber sido tan grande como en los Hominini anteriores.
- Son de mayor tamaño que los Hominini anteriores y maduraban lentamente. En el nacimiento eran muy inmaduros (como los niños actuales) debido al compromiso entre el tamaño de la cabeza y el del canal del parto.
 |
| Significado de KNM-WT 51260. Crédito: Adam Benton |
Para Wood y Collard (1999) habilis/rudolfensis, presentan un tamaño corporal mejor adaptado que el erectus a un ecosistema cerrado. El incremento del tamaño corporal en erectus mejora la termorregulación en entornos más áridos. Sus caracteres indican el abandono de toda condición arborícola y una locomoción prácticamente idéntica a la de los humanos modernos:
- Grandes superficies articulares en los huesos largos.
- Gran espesor cortical en los huesos de las piernas.
- Pelvis estrecha con pilares ilíacos marcados.
- Extremidades superiores más cortas.
El tiempo de la muda de los dientes se va alargando lo que puede indicar una prolongación de la inmadurez y del tiempo de aprendizaje.
Diferencias entre erectus africanos y asiáticos
Se ha especulado con una separación entre los ejemplares africanos y los asiáticos. Por ejemplo, Wood (1984) describe las siguientes autapomorfias para los erectus asíaticos:
- Torus occipital con sulcus superior.
- Torus angular y cresta mastoidea.
- Toro supraorbital separado de la escama frontal por el surco posttoral.
- Proporciones y forma del hueso occipital.
- Arco occipital relativamente largo.
Para J. L. Arsuaga, los erectus asiáticos presentan superestructuras craneales mucho más marcadas:
- Torus frontal muy desarrollado y recto.
- Hueso occipital más angulado. Torus occipital muy marcado.
- Mayor grosor de las paredes del cráneo y de los huesos del esqueleto.
- Bóveda craneal baja.
- Frente huidiza.
- Base del cráneo ancha.
- Quilla sagital (borde longitudinal en V a lo largo de la parte superior del cráneo). No se conoce la función de esta quilla, pues no servía para insertar músculos temporales, como en los parántropos.
Te puede interesar:
|
Homo ergaster y Homo erectus
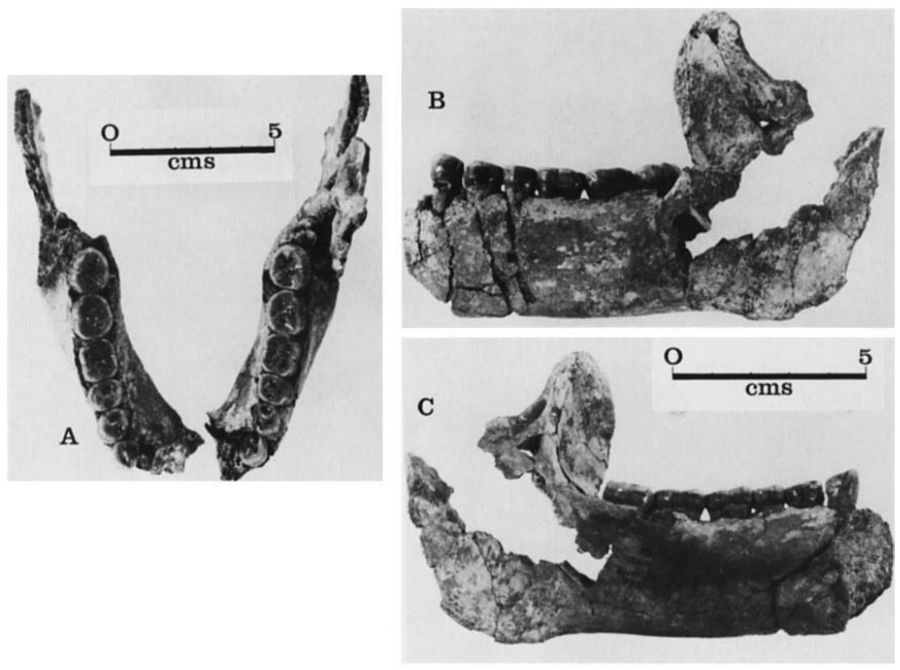 |
| KNM-ER 992. Leakey y Wood, 1973. |
- Gagneux (2011) mantiene que hace 3 Ma, los Hominini portadores de una variante genética que inactivó el correspondiente gen empezaron a producir la variante del ácido siálico Neu5Ac, que es más resistente a los parásitos. Estos seres desarrollaron una respuesta inmune a la variante presente en los monos antropomorfos y en los Hominini ancestrales (Neu5Gc) que afectó a la reproducción ya que los anticuerpos matan el esperma con Neu5Gc. Según Xiaoxia Wanga et al (2012) la eliminación genética de los receptores del ácido siálico Siglec-13 y / o Siglec 17 aumentó la resistencia a las enfermedades infecciosas y a los procesos inflamatorios y contribuyó a eliminar las restricciones al crecimiento de las poblaciones. La resistencia a las enfermedades junto a este aislamiento reproductivo pudo facilitar la divergencia de este grupo hasta constituir el grado erectus.
El ergaster migró rápidamente a Asia (1,8-1,7 Ma), dando lugar a Homo erectus. Los fósiles africanos DNH 134, un fragmento de cráneo, y KNM-ER 2598, un fragmento occipital, son anteriores a los fósiles de Dmanisi y Java, lo que indica que comenzaron en África antes de hace 1,9 Ma y después emigraron al continente Euroasiático. En el sudeste de Asia, Homo erectus persistió hasta el último Pleistoceno. Homo ergaster persiste en África durante más de un millón de años, hasta hace 0,78 Ma. Por consiguiente, estos taxones se mantienen en estasis durante centenares de miles de años. Tras la década de 1950, los hallazgos más importantes se han producido en África.
Durante miles de años, el cerebro sigue creciendo lentamente hasta alcanzar 1.100 cc en los últimos ejemplares. Junto con el aumento de cerebro, se incrementan también las capacidades cognitivas: en sus comienzos, los ergaster mantienen las condiciones culturales olduvaienses de los habilis. Más tarde se producen adelantos significativos en la fabricación de herramientas que dan lugar a una nueva tradición cultural: la achelense, ausente en Asia, con alguna excepción, y que no aparece en Europa hasta época tardía (0,5 Ma tras una oleada moderna).
| El sendero evolutivo hacie el erectus. |
Hasta la década de los 50, erectus se definía de una forma temporal, entre el final de los australopitecinos sudafricanos y el comienzo de los sapiens arcaicos de Europa. A medida que se fueron encontrando autapomorfias, se afirmaba su imagen como un ser característico pero se debilitaban las posibilidades de relacionarlo con sus antecesores (Homo habilis) y sucesores.
- Homo erectus, Dubois, 1892, 1894; Weidenreich, 1940. Hombre erecto. Incluye los hallazgos de Dubois en Java a finales del XIX y los de China en la década de 1920. Weindenreich estableció los criterios diagnósticos en las décadas de 1930 y 1940.
- En 1950, Ernst Mayr propuso agrupar la mayoría de los fósiles atribuidos al género Homo conocidos hasta entonces en Homo erectus. A medida que los nuevos hallazgos se han ido acumulando, para los taxónomos agrupacionistas el grado erectus sensu lato ha ido reuniendo ejemplares tan diferentes como el Homo georgicus o el Homo floresiensis.
- Homo ergaster, Groves y Mazák, 1975. Hombre trabajador.
- En todo caso, tuvieron un ancestro común próximo
Para muchos investigadores sus grandes similitudes y la dificultad de asignar algunos restos a uno de los dos taxones recomiendan considerar estas especies en conjunto formando lo que podemos llamar el grado erectus siguiendo el concepto grado de Julian Huxley (1958; unidad de progreso anagenético).
La postura que se adopte depende de la forma de entender lo que fue la evolución humana:
- bien como un proceso un tanto conservador, con ciertas modificaciones locales que no llegaron a dividir las poblaciones en varios especies.
- o como una diversidad de líneas que dieron lugar a ciertas especiaciones geográficamente dispersas (Homo erectus en Asia, los neandertales en Europa, Homo ergaster y los primeros seres humanos de aspecto moderno en África).
- Esta última debe resolver un problema práctico. Los Homo del Pleistoceno Medio manifiestan sin lugar a dudas una cierta unidad. Son un mismo “tipo” de Hominini que pueden relacionarse con unos caracteres anatómicos y unos rasgos de conducta muy precisos. Si los separamos en dos o más especies ¿cómo cabe expresar esa unidad? Cualquier suma de Homo erectus sensu stricto y de Homo ergaster constituirá un grupo parafilético, porque –si se acepta el esquema diversificador- el segundo forma parte del linaje de Homo sapiens mientras que el primero constituye un clado lateral, su grupo hermano.
 |
| Posturas acerca de la clasificación de los Hominini del Pleistoceno Inferior y Medio |
Reducción a una sola especie estable morfológicamente (hipótesis de la estasis)
- Hipótesis de la especie única (erectus), defendida por R. Leakey y Walker (1976), basándose en que en África aparecieron restos parecidos a los asiáticos y que no podía haber más de una especie Hominini en el mismo lugar y tiempo.
- Howells (1980) y Rightmire (1981, 1990) entre otros, consideran que los especímenes atribuidos a erectus permanecieron sensiblemente iguales a lo largo del tiempo sin cambios morfológicos notables y dieron lugar a sapiens a través de una transición brusca (equilibrio puntuado).
- Bräuer (1994) sostiene que los dimorfismos sexuales y la alta variabilidad regional son suficientes para explicar la diversidad de los erectus de África y Asia y que en consecuencia todos ellos deben ser atribuidos a la misma especie.
- Para Wolpoff (1999) la continuidad ha sido tan acusada que solo cabría hablar de una única especie de Homo, a la que pertenecemos nosotros. Defiende cambios pequeños y continuos.
- Wood y Collard (1999), siguiendo el criterio de Huxley, agrupan erectus y ergaster en el grado erectus, caracterizado por un aumento notable de la capacidad craneal, aunque matizado por un aumento notable del tamaño del cuerpo. Hubo presiones selectivas, a las que los Hominini respondieron aumentado de tamaño. Las técnicas de construcción de útiles de piedra fueron ganando en complejidad, lo que lleva a entender que las capacidades cognitivas se incrementaron notablemente. Esta es la tesis por la que se inclinan Camilo José Cela Conde y Francisco Ayala.
- Emiliano Bruner et al (2014) han analizado la variación endocraneal de especímenes africanos y asiáticos pertenecientes a la hypodigma de H. erectus sensu lato, para evaluar si se pueden definir distintos grupos regionales. De acuerdo con los resultados, los ejemplares de China, Indonesia y África se superponen en gran medida en su distribución fenotípica y patrones morfológicos.
- Igulamente, Baab (2016) concluye que los erectus de China, Indonesia, Georgia y el este de África comparten una forma neurocraneal distinta de la de otros taxones del Plio-Pleistoceno. En la variación morfológica del erectus, los ejemplares de Indonesia forman una puente entre las poblaciones anteriores y posteriores de África y Asia.
Varias especies
- Los investigadores que distiguen H. ergaster de H. erectus, consideran que el primero es más primitivo:
- Menor tamaño de los premolares y sobre todo de los molares.
- Bóveda craneal menos especializada.
- Esta propuesta no ha ganado un reconocimiento general, pero es un buen medio para reconocer que los erectus africanos y asiáticos no pertenecen a la misma especie. El aislamiento y la deriva genética habrían dado lugar a una especie diferenciada.
- Sus culturas no coinciden.
- Las dataciones no corresponden.
- Tendríamos ergaster en África y erectus en Asia.
- El erectus sería entonces una especie asiática lateral a la nuestra, si bien Groves consideró que el ejemplar OH9 de Olduvai (1,2 Ma) constituye una subespecie que representa la única presencia del erectus en África.
- Homo sería un grupo parafilético, pero compartiendo unos caracteres anatómicos y unos rasgos de conducta muy precisos.
- Los dientes de los erectus asiáticos exhiben una serie características exclusivas: unas arrugas muy marcadas en la dentina y una gran robustez con raíces masivas (Xing et al, 2018)
Posición intermedia
 |
| Hallazgos de Homo ergaster y erectus y dataciones |
Te puede interesar:
|
viernes, 27 de septiembre de 2013
Estudio sobre los restos neandertales hallados en la Cueva Kalamakia.
 |
| Molares hallados en Kalamkia. |
Inventario de los restos humanos de Kalamakia.
miércoles, 25 de septiembre de 2013
La expansión de la población produjo el Neolítico y no al contrario.
C. Aimé, G. Laval, E. Patin, P. Verdu, L. Ségurel, R. Chaix, T. Hegay, L. Quintana-Murci, E. Heyer y F. Austerlitz analizaron el ADN nuclear (404 individuos de 16 poblaciones) y mitocondrial (2429 de 61) de poblaciones humanas de África y Eurasia con la finalidad de inferir su historia demográfica a partir de las huellas en el poliformismo genético. Compararon los resultados genéticos con los hallazgos arqueológicos.
Según sus conclusiones:
Según sus conclusiones:
- En África, se produjeron eventos de expansión relacionados con la agricultura, mientras que las poblaciones de cazadores recolectores permanecieron constantes o se contrajeron.
- Una expansión de acuerdo con el modelo de expansión para Chaggas y Mozambiqueños (125-62 ka).
- Una expansión de acuerdo con el modelo exponencial para Akeles, Ngumbas y Yorubas (16-8 ka).
- Una expansión en las poblaciones actualmente agrícolas de África Central entre 91/45-63/31 ka.
- Una expansión en las poblaciones agrícolas entre 5-4 ka.
- En Eurasia, se infieren tasas de expansión más altas para los agricultores que para los pastores excepto en Asia Central y Corea.
- Una expansión de acuerdo con el modelo exponencial para los chinos Han (72-36 ka).
- Para Japonenses, Daneses y Chuvash se aprecia una contracción o población constante.
- En Asia Central, todas las poblaciones muestran expansión. Los actuales agricultores desde hace 33 o 17 ka. Los actuales pastores desde hace 28 o 14 ka.
- En resumidas cuentas, las expansiones de población podrían haber comenzado en el Paleolítico (80-60 ka) quizá gracias a nuevas tecnologías aplicadas a la caza o a un cambio ambiental rápido hacia climas más secos, favoreciendo la orientación de las correspondientes poblaciones hacia una agricultura durante el Neolítico. Las poblaciones que adoptaron un estilo de vida agrícola sedentario durante el Neolítico habían experimentado previamente las expansiones más fuertes durante el Paleolítico.
lunes, 23 de septiembre de 2013
Se confirma la utilidad del foramen magnum como indicador de la bipedación
 |
| Posición del foramen magnum en hominoideos |
Gabrielle A. Russo y E. Christopher Kirk han estudiado la posición del foramen magnum en especies bípedas y cuadrúpedas de tres clados mamíferos: marsupiales, roedores y primates. Además, examinaron si los primates estrepsirrinos que habitualmente asumen posturas troncales ortógradas, exhiben unos foramen magna posicionados más anteriormente en comparación con estrepsirrinos no ortógrados.
Según las conclusiones, los marsupiales y roedores bípedos muestran foramen magna en posiciones más anteriores que en sus parientes cercanos cuadrúpedos. El foramen magnum se encuentra también más anterior en estrepsirrinos ortógrados que en estrepsirrinos pronógrados o antipronógrados. Entre los primates incluidos en la muestra, los seres humanos presentan los foramen magna en una posición más anterior.
Los resultados de este análisis apoyan la utilidad de la posición del foramen magnum como un indicador de la locomoción bípeda en los homínidos fósiles.
Enlace
Reseña de la Universidad de Texas
Conclusiones contrarias en Ruth et al (2016)
jueves, 19 de septiembre de 2013
Los orangutanes, planifican el futuro
 |
| Debido a que sus adiposidades y bolsa gular actúan como un megáfono, el sonido de las llamadas de larga distancia se transmite direccionalmete, y de esta forma las hembras pueden conocer la futura ruta del macho, en la misma dirección que la llamada. Si la hembra está en la posición B, no necesitará desplazarse mucho, solo salir al encuentro del macho. |
Los orangutanes macho maduran alrededor de los 15 años de edad, momento en el cual tienen testículos completamente descendidos y pueden reproducirse. Entre los 15-20 años, en ausencia de un macho residente, desarrollan unas adiposidades distintivas en la mejilla, bolsas pronunciadas en la garganta (bolsa gular) y pelo largo y efectúan llamadas de larga distancia. El desarrollo de estas características depende en gran medida de la ausencia de un macho residente. Los machos sin ellas se conocen como machos unflanged, mientras que los que las poseen se denominan flanged. Estos distintos tipos de macho tienen estrategias de apareamiento diferentes. Los machos flanged atraen a las hembras con sus características llamadas de larga distancia. Esas llamadas pueden suprimir el desarrollo en los machos más jóvenes. Los machos unflanged vagan en busca de hembras en celo y las obligan a la cópula cuando las encuentran. Ambas estrategias tienen éxito, pero las hembras prefieren aparearse con machos flanged y buscan su protección contra los unflanged. Estas asociaciones macho-hembra son estables.
Carel P. van Schaik, Laura Damerius y Karin Isler han comprobado en los orangutanes macho de Sumatra (Pongo abelii) que la dirección en la que emiten sus llamadas predicen su posterior dirección de marcha con mucha antelación, y que una nueva llamada indica un cambio de dirección. Estos resultados muestran que los orangutanes macho hacen planes de viaje y los anuncian a sus congéneres. Esta capacidad de planificar el futuro se pensaba que era únicamente humana.
Orangutanes
lunes, 16 de septiembre de 2013
Nuevas dataciones del musteriense y el achelo-yabrudiense de Monte Carmelo.
 |
| Situación de la Cueva Misliya |
- Las edades medias obtenidas para las muestras asociadas al Achelo-Yabrudiense (reforzadas por un análisis isócrono) indican una producción de este complejo cultural desde el fiunal de MIS 8 hasta hace ~ 250 ka.
- Para las industrias del Paleolítico Medio Temprano, ricas en productos Levallois y laminares se asignaron edades medias que van de ~ 250 a ~ 160 ka (MIS 7 y primera parte de MIS 6).
- El paso del Paleolítico Inferior al Medio tuvo lugar en el límite MIS 8/7 o durante la primera parte de MIS 7. Las fechas, junto con las marcadas diferencias en la tecnología lítica sugieren fuertemente la llegada de una nueva población durante este período.
Asia Occidental: donde se juntan los caminos y las especies
jueves, 12 de septiembre de 2013
Nuevas dataciones de Ksar Akil cuestionan la procedencia de los europeos modernos
 |
| Estratigrafía de Ksar Akil |
Katerina Douka, Christopher A. Bergman, Robert E. M. Hedges, Frank P. Wesselingh, y Thomas F. G. Higham han datado por radiocarbono los contextos donde fueron hallados dos fósiles de HAM en Ksar Akil (Líbano) y los niveles superior e inferior:
- Egbert, un esqueleto juvenil completo, actualmente extraviado y
- Ethelruda, un maxilar redescubierto recientemente y que fue clasificado en un principio como “neandertaloide”.
Para la datación, los autores utilizaron principalmente cuentas de concha marina, consideradas como un indicio del comportamiento moderno. El modelo estimó unas dataciones entre 40,8–39,2 ka para el lugar donde fue hallado Egbert y 42,4–41,7 ka para el de Ethelruda. Por consiguiente, Egbert tiene una antigüedad comparable a la de los restos de HAM de Peştera cu Oase y Ethelruda a la de los dientes de HAM de Cavallo.
La datación de las capas del Paleolítico Superior Temprano indican que el paso desde el Paleolítico Medio al Superior en Ksar Akil, y posiblemente en toda el área geográfica sucedió más tarde de lo que se pensaba, lo que arroja algunas dudas sobre el papel de la región como un locus para la dispersión humana a Europa. La datación e interpretación de los fósiles del contexto de Ethelruda son un indicio de que las industrias transicionales o del Paleolítico Superior Inicial (IUP) en Europa y el sur de Asia Occidental, o al menos algunas de ellas, son resultado de migraciones tempranas del HAM.
Enlace
Comentarios de Sarah Griffiths
Comentarios en Science Daily
Comentarios en Past Horizons
Comentarios en Hominidés
Comentarios de Guillermo Caso de los Cobos
La datación de las capas del Paleolítico Superior Temprano indican que el paso desde el Paleolítico Medio al Superior en Ksar Akil, y posiblemente en toda el área geográfica sucedió más tarde de lo que se pensaba, lo que arroja algunas dudas sobre el papel de la región como un locus para la dispersión humana a Europa. La datación e interpretación de los fósiles del contexto de Ethelruda son un indicio de que las industrias transicionales o del Paleolítico Superior Inicial (IUP) en Europa y el sur de Asia Occidental, o al menos algunas de ellas, son resultado de migraciones tempranas del HAM.
Enlace
Comentarios de Sarah Griffiths
Comentarios en Science Daily
Comentarios en Past Horizons
Comentarios en Hominidés
Comentarios de Guillermo Caso de los Cobos
Asia Occidental. Donde se juntan los caminos y las especies
 |
| Datación de varios yacimientos paleolíticos. |