- Reducción en la importancia relativa del olfato.
- Incremento en la importancia relativa de la visión.
- Con excepción de los monos búho de Sudamérica, todos los antropoides son de actividad diurna. Debido a su agudeza visual y visión de color, los primates pueden detectar fácilmente la coloración de una fruta madura, o la hinchazón de color rojo de un potencial compañero, así que dependen de la información visual para tener éxito en su entorno. La capacidad de ver en tres dimensiones es un rasgo de la gran capacidad de adaptación, pues los primates pasaron gran parte de su historia evolutiva en los árboles. El aumento relativo en el tamaño y la complejidad de la corteza visual en los primates, y particularmente en los antropoides, es consecuencia de este cambio adaptativo.
- Enorme incremento en la importancia absoluta y relativa de la neocorteza.
- La neocorteza está involucrada en el procesamiento de la información. Durante la evolución del cerebro antropoide, los primates con mayores áreas de asociación para procesar la información tenían más probabilidades de transmitir sus genes a la siguiente generación. Eran más listos, tenían mayor flexibilidad de comportamiento y más capacidad de adaptarse a los cambios del entorno.
 |
| Comparación mediante escalas logarítmicas entre el peso corporal en gramos y el peso del cerebro en miligramos en 45 especies de primates. La línea (regresión) muestra el tamaño cerebral esperado para cada tamaño corporal. La X sobre la línea de regresión marca el tamaño esperado para el cerebro humano, lo que resulta mucho menor que el verdadero tamaño del cerebro humano actual, señalado con un cuadrado de líneas rojas. En general, los prosimios se ubican bajo la línea de regresión, en tanto que los antropoides se ubican sobre esta línea. Esta característica indica que hubo una presión selectiva para el desarrollo del tejido cerebral durante la evolución humana. Rosales-Reynoso et al, 2018. Tomado y modificado de Stephan et al, 1981. |
- La hipótesis del radiador (Falk, 1990) sugiere que la expansión del cerebro en Homo fue posible gracias a una reconfiguración del suministro vascular craneal en respuesta a los cambios en la presión hidrostática producidos por la bipedación.
- La hipótesis del crecimiento lineal (Finlay y Darlington, 1995) considera que los componentes cerebrales no aumentaron de tamaño en mosaico sino de forma uniforme y concertada, de acuerdo con sus propias reglas internas y con una escala alométrica predecible.
- La hipótesis del tejido costoso (Aiello y Wheeler, 1995, 2002), mantiene que el aumento de tamaño del cerebro es caro con relación al resto del cuerpo en términos de energía y ello limita el aumento de tamaño. En los orígenes humanos, se produjo un importante cambio en la asignación de la energía con el fin de apoyar la notable expansión de un cerebro metabólicamente costoso.
- Fedrigo et al (2011) han comprobado que la expresión de los genes SLC2A1 y SLC2A4 que regulan el transporte de glucosa es diferente en chimpancés y humanos. En aquellos son más activos en los músculos mientras que en los humanos son más activos en el cerebro. En los humanos, estos genes muestran firmas de selección positiva.
- La hipótesis de la energía materna (Martin, 1996) propone que la tasa metabólica relativa de las madres durante la gestación de los bebés determina la masa cerebral neonatal del recién nacido, mientras que después nacimiento, la inversión materna de las madres durante el crecimiento de su descendencia podría ser otro de los factores implicados en el desarrollo del cerebro.
- La hipótesis del cerebro social (Dunbar y Shultz, 2007; Shultz y Dunbar, 2007) postula que fueron las demandas computacionales de la vida en grandes sociedades complejas las que produjeron la selección de grandes cerebros.
- La hipótesis de la complejidad del entorno sugiere que la función de la cognición es permitir al organismo adaptarse a entornos heterogéneos (Godfrey-Smith, 2002; Grove, 2017).
 |
| Evolución de la capacidad craneal. Miriam Noël Haidle (2012) |
ALOMETRÍA.
A lo
largo de la evolución humana se ha manifestado una tendencia al aumento del
volumen corporal.
Un
cuerpo más voluminoso es más eficiente energéticamente. En el metabolismo de
los seres vivos, una secuencia repetida de reacciones químicas descompone la
materia orgánica, la transforma en energía y la pone en uso para el
mantenimiento y reproducción celular. La tasa metabólica de un organismo es
igual a la cantidad de energía necesaria para mantenerlo vivo.
Pensemos
en un ratón como una caja de 10x5x5 cm y en un elefante como una de 4x3x2 m.
Ambos están constituidos por células que producen metabolismo y liberan calor
como subproducto. El calor se disipa a través de la superficie. El ratón de
nuestro ejemplo tiene una superficie de 250 cm2 y un volumen de 250
cm3. El elefante, una superficie de 52 m2 y un volumen de
24 m3. El elefante tiene una superficie 2.080 veces superior a la
del ratón y un volumen 96.000 veces mayor. Debido a que el tamaño celular es
similar en todos los mamíferos, el elefante está formado por 96.000 veces más
células que el ratón, pero dispone de una superficie relativamente menor para
disipar el calor del metabolismo. Por consiguiente, el metabolismo del elefante
ha de ser mucho más lento. En el mismo periodo de tiempo consume muchos menos
recursos por célula.
En
definitiva, se puede conseguir un aumento corporal apreciable con un pequeño
incremento de consumo de recursos. Por esta razón, un aumento de tamaño puede
ser una buena estrategia evolutiva cuando los recursos abundan. Un mayor tamaño
puede proporcionar un mejor acceso a los recursos y mejores resultados en el
ataque y defensa. El tamaño puede ser además objeto de selección sexual.
Cuando
el cuerpo aumenta de volumen en forma proporcional, hablamos de una expansión isométrica. Si una parte del cuerpo aumenta más que proporcionalmente decimos
que se da una expansión alométrica. Este es el caso del cerebro durante la
evolución humana, con un aumento mayor del esperado.
 |
| Porcentaje del volumen cerebral dedicado al bulbo olfatorio, la corteza visual y la neocorteza en prosimios, monos, simios y humanos actuales. Rosales-Reynoso et al, 2018. Tomado y modificado de Stephan et al, 1981. |
- Durante la evolución de Homo, aumentó el volumen del cerebro y disminuyó el tamaño de los dientes poscaninos (con alguna excepción, p.ej. Homo floresiensis).
- Por el contrario, en el resto de los primates vivos, salvo los platirrinos, el tamaño de los poscaninos y el cerebro muestran ambos una alometría negativa.
- Los australopitecos muestran pequeñas variaciones a lo largo del tiempo tanto en el tamaño de los dientes como en el volumen del cerebro.
- En los parántropos los dientes aumentan de tamaño (megadoncia) mientras que la masa corporal y el tamaño del cerebro no aumentan significativamente.
 |
| Relación entre el volumen del cerebro (BrM) y el tamaño de los dientes poscaninos (PCTA). La regresión de Homo se calcula sin tener en cuenta a Homo floresiensis. Juan Manuel Jiménez-Arenas et al (2014). |
Yue Huang et al (2013) han verificado que los genes objeto de selección positiva que se expresan en el sistema central, especialmente los relacionados con las funciones cognitivas, han sido los últimos en evolucionar.
Para André Leroi-Gourhan (1964), el desarrollo temprano de la bipedación no solo liberó las manos para las acciones técnicas; el cambio de posición del foramen magnum permitió un aumento de tamaño de los lóbulos occipitales. Posteriores soluciones técnicas sencillas, como las primitivas herramientas de piedra, permitieron la reducción del aparato masticatorio, lo que posibilitó cambios en el parietal y en especial en los lóbulos frontales. La boca, dedicada menos tiempo a la nutrición, pudo usarse para la comunicación vocal. El factor sociocultural se hizo cada vez más importante desde el final del Paleolítico Medio, cuando el grupo étnico actuó como un banco de memoria social y el lenguaje se utilizó para el almacenamiento cultural. El resultado fue la explosión de las variedades de artefactos y símbolos del Paleolítico Superior. La evolución física del cerebro permitió la emergencia de nuevas capacidades para el pensamiento.
Para André Leroi-Gourhan (1964), el desarrollo temprano de la bipedación no solo liberó las manos para las acciones técnicas; el cambio de posición del foramen magnum permitió un aumento de tamaño de los lóbulos occipitales. Posteriores soluciones técnicas sencillas, como las primitivas herramientas de piedra, permitieron la reducción del aparato masticatorio, lo que posibilitó cambios en el parietal y en especial en los lóbulos frontales. La boca, dedicada menos tiempo a la nutrición, pudo usarse para la comunicación vocal. El factor sociocultural se hizo cada vez más importante desde el final del Paleolítico Medio, cuando el grupo étnico actuó como un banco de memoria social y el lenguaje se utilizó para el almacenamiento cultural. El resultado fue la explosión de las variedades de artefactos y símbolos del Paleolítico Superior. La evolución física del cerebro permitió la emergencia de nuevas capacidades para el pensamiento.
 |
| Modelo de Leroi-Gourgham de la evolución cognitiva. |
 |
La evolución del cerebro
La evolución del cerebro se ha caracterizado por:
- Un aumento extraalométrico de la capacidad craneal (pero Grabowski, 2016).
- Los primeros homínidos tienen unas capacidades craneales semejantes a las de los chimpancés actuales. Éstos son capaces de utilizar herramientas simples como palos o piedras e incluso establecen distintas tradiciones de manipulación aprendida de objetos, pero no parecen disponer de la capacidad de anticipación, planificación y coordinación necesarias para llevar a cabo la cultura olduvaiense.
- El incremento de la capacidad craneal puede ser un buen indicio acerca de la aparición de capacidades cognitivas nuevas (Sarah Benson-Amram et al, 2016), sobre todo si tenemos en cuenta lo caro que es, en términos metabólicos, un órgano como el cerebro. Además, un aumento de tamaño permite un aumento de complejidad y mayor capacidad de cálculo (Teoría Computacional de la Mente). Esta medida suele bastar cuando se comparan animales del mismo grupo filogenético. Pero los antropólogos utilizan también procedimientos estadísticos.
- El coeficiente de encefalización mide la relación existente entre el peso del cerebro y el peso del cuerpo).
- Cuando comparamos animales de distintos grupos filogenéticos, la exhibición de habilidades intelectuales está relacionada con la proporción alométrica entre el volumen del encéfalo y el tamaño de los animales. Se supone que el tamaño del cuerpo implica para su gestión un tamaño cerebral determinado y el exceso supone una capacidad computacional adicional.
- Entre los monos del Nuevo Mundo, los que tienen alimentación insectívora cuentan con mayor coeficiente que los que se alimentan de hojas y ese hecho se relaciona con la mayor necesidad de procesamiento de la información procedente del medio ambiente.
- Harry Jerison efectuó un ajuste mediante regresión lineal entre los logaritmos de la masas corporal y cerebral de los vertebrados. La pendiente resultó ser aproximadamente 2/3, y este dato permite estimar el volumen cerebral esperado de cualquiera de estas especies. Los humanos tenemos un encéfalo casi ocho veces mayor de lo esperado (tres veces si solo tenemos e cuenta los primates antropoideos).
- La relación entre el tamaño cerebral en peso y la superficie corporal en área.
- Un aumento en la superficie del córtex.
- La relación entre la extensión del neocórtex y el volumen de la médula espinal (Passihgham, 1975). Se apunta de manera muy generalizada a una expansión del córtex frontal (Deacon, 1997), especialmente algunas regiones muy anteriores, como el área 10 de Brodmann. En los humanos abundan las células Von Econo, especialmente conectadas con el área 10 y asociadas a las relaciones sociales.
- La girificación es la relación que existe, en cualquier sección del córtex, entre la longitud del perímetro cortical exterior (trazando el contorno más externo del córtex sin seguir sus recovecos) y la del perímetro total (midiendo el interior también de los plegamientos que hacen que el córtex humano parezca algo similar a una coliflor, salvadas sean todas las distancias).
- Herculano-Houzel ha llegado a la conclusión de que los primates evolucionaron empacando muchas más neuronas en la corteza cerebral que otros mamíferos. Los seres humanos parecen disfrutar de más neuronas corticales que cualquier otra especie.
- El área de la superficie externa del cerebro, lo que corresponde aproximadamente a la superficie endocraneal, crece con un exponente alométrico de ~ 2/3 en relación con el volumen del cerebro. Sin embargo, la materia gris, que constituye la capa más externa del cerebro y contiene la mayoría de las neuronas, aumenta casi linealmente con el aumento de volumen del cerebro (exponente 0,955). Esto se traduce en un problema de embalaje espacial, que se resuelve mediante una intensificación del plegado cortical (girification). El gran número de neuronas también crea una "sobrecarga de comunicación," sustancial a medida que más neuronas necesitan estar interconectadas a través de distancias más grandes. La materia blanca, que constituye los haces de axones mielinizados por los que las regiones se comunican entre sí, aumenta a un exponente de 1,233. En los primates, este exponente es algo más pequeño (1,148). La girificacion es uno de los principios de construcción por el que las distancias de comunicación se mantienen cortas. Debido a que los grandes cerebros imponen limitaciones específicas de la organización neuroarquitectural, la ampliación de la evolución del cerebro de los homínidos siempre implica reorganización.
- El gen TRNP1 parece ser el responsable de la producción de pliegues, ya que se expresa en niveles bajos en las zonas con pliegues y en niveles altos en las zonas más lisas. Otro gen implicado en la formación de neuronas y pliegues puede ser el HARE5.
- Marta Florio et al (2015) han analizado los transcriptomas en ratones y humanos, identificando 56 genes que se expresan preferentemente en la glía radial apical y basal de los humanos, células madre de la corteza cerebral, y que carecen de ortólogos en el ratón. Entre estos, ARHGAP11B tiene el más alto grado de expresión. ARHGAP11B surgió de la duplicación parcial del gen ARHGAP11A codificador de las proteínas de activación de las GTPasas Rho, que regulan un alto número de procesos de señalización y estructurales de la célula en respuesta a diversos estímulos. Esta duplicación se produjo después de la separación con los chimpancés. ARHGAP11B solo está presente en el genoma del humano (HAM, neandertales y denisovanos) y produce la formación de los pliegues cerebrales (aumento de la girificación) (Marta Florio et al, 2017). Los autores analizaron la función de este gen en el desarrollo del neocórtex y en la generación de neuronas desde las células progenitoras mediante un experimento con ratones. Insertaron el gen ARHGAP11B en el cerebro embrionario de un ratón, cuyo neocórtex es completamente liso y cuenta con un número muy pequeño de células madre. La inserción del gen en el cerebro en desarrollo del ratón provocó que el número de células progenitoras de la corteza aumentara hasta llegar casi al doble y el neocórtex comenzó a arrugarse.
- Es posible una evolución en el sentido de una mayor girifación tal como sucedió en Homo, pero también en el sentido contrario, como parece haber sucedido en los monos del Nuevo Mundo.
- Boldog et al (2018) han reportado la existencia de un subtipo específico humano de neuronas GABAérgicas.
- Mayor diversidad de áreas corticales y superficies más desarrolladas en los lóbulos parietales.
- El surco intraparietal es mucho más grande y complejo que en los otros primates. Esta implicado en la relación ojo-mano.
- El precúneo está mucho más desarrollado que en los chimpancés, en particular las áreas involucradas en la relación cuerpo-visión y cuerpo-espacio, la imaginación y la autoconciencia.
- Aparición del opérculo frontal, dentro del que se observan varias estructuras que creemos comprometidas con el lenguaje como el área de Broca, el comportamiento social y la planificación motriz.
- Homo naledi, la población de la Sima de los Huesos en España e incluso en algunos Homo erectus (hace 1,5 Ma) muestran opérculo frontal, mientras que está ausente en los ejemplares anteriores a hace 1,8 Ma (Homo rudolfensis, Homo habilis y Australopithecinos).
- Mayor tamaño de los lóbulos temporales, en relación al patrón de los grandes simios.
- Estas áreas están involucradas en el lenguaje y la memoria.
- Menor tamaño de los lóbulos occipitales, en relación al patrón de los grandes simios.
- Relacionados con la decodificación de las señales visuales.
- Una mejora de la conectividad neuronal.
- Schoenemann, Sheehan y Glotzer (2005) midieron el volumen de la materia gris, blanca y el volumen total del lóbulo prefrontal y de todo el cerebro en 11 especies de primates. En términos relativos, se encontró que la materia blanca (conexiones de las neuronas con mielina) del lóbulo prefrontal, fue la mayor diferencia entre los humanos y los no humanos, mientras que la materia gris no mostró diferencias significativas.
- Están particularmente potenciadas la vías que unen los centros del lenguaje y las conexiones entre las áreas frontales y parietales.
- Cambios en los neurotransmisores.
- Los homínidos muestran un mayor contenido de serotonina y dopamina que el resto de los primates. La acetilcolina, que participa en la regulación del comportamiento jerárquico y territorial, se encuentra menos en los humanos que en los gorilas y chimpancés (Raghanti et al, 2018).
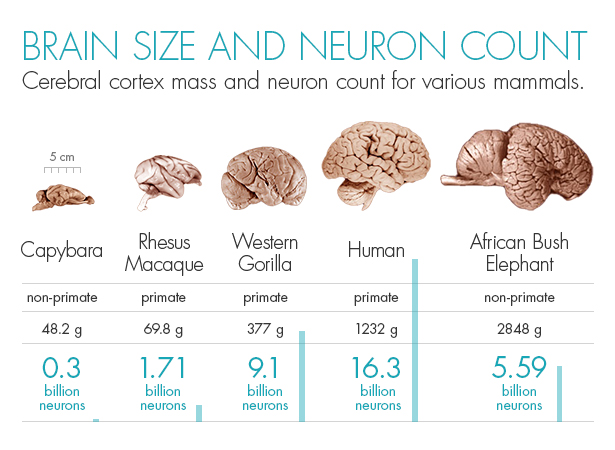 |
| Masa cerebral y número de neuronas en algunos mamíferos. Olena Shmahalo / Revista Quanta; fuente: BrainMuseum.org y Herculano-Houzel et al. |
Etapas en la evolución del cerebro.
Según Aida Gómez Robles, William D. Hopkins y Chet C. Sherwood (2014) el cerebro está dividido en módulos poco integrados, lo que pudo favorecer su evolución en mosaico.
Etapas:
- Reorganización del parietal posterior y del occipital anterior hacia pautas más humanas, pero sin cambios apreciables por lo que hace al volumen cerebral, que continúa en Australopithecus dentro de los valores de los simios.
- En las primeras etapas de la evolución de los hominini se produjo una reorganización vascular y cortical.
- Drenaje de la sangre a través de los senos marginales occipitales agrandados a expensas del sistema de seno transverso / sigmoide en los australopitecinos robustos.
- Aumento de ramificación de la arteria meníngea media en los homínidos de gran cerebro.
- Robert Martin (2000) supone una escasa diferenciación sexual en el Australopithecus africanus y por ello cree que se sobreestimó su tamaño corporal, de modo que el incremento de encefalización se daría ya en esta especie, posiblemente relacionado con la actividad locomotora y la búsqueda de un tipo diferente de alimentos a su vez relacionados con las necesidades de un cerebro en expansión.
- Falk et al (2000) comparando los endocráneos de diferentes especies de parántropos y los de Austraopithecus africanus dan una idea de la evolución cerebral parecida a la de Martin, apoyando la hipótesis del Australopithecus africanus como un ancestro del Homo.
- Los endomoldes de Australopithecus africanus indican una reorganización del cerebro con un aumento sustancial en el tamaño (aunque esa reorganización probablemente no tuvo lugar en el Paranthropus). Los polos frontales son cuadrados, y los polos temporales se ampliaron anterolateralmente, lo que indica la expansión de los lóbulos frontal y temporal.
- Hay evidencia de que australopitecinos dieron a luz a bebés comparativamente de gran cerebro, lo que implica modificación evolutiva del desarrollo perinatal.
- Según Falk et al (2000) y Falk et al 2012, el endocasto de Taung 1 reproduce una sutura metópica parcialmente fundida. Esta condición es relativamente frecuente en los jóvenes adultos humanos modernos, pero rara en los chimpancés. La fusión retardada de la sutura metópica podría ser adaptativa en varios aspectos: facilita el nacimiento de bebés de cerebro grande y permite la rápida expansión postnatal del cerebro, en especial de la corteza frontal. Ralph L. Holloway, Douglas C. Broadfield y Kristian J. Carlson (2014) han utilizado tomografía computerizada de imágenes de alta resolución de Taung 1 y no han observado fusión retardada de la sutura metópica ni variación de tamaño de la región prefrontal con respecto al chimpancé.
- El hecho de naledi muestre unas estructuras neurológicas equivalentes a sapiens pero con un tamaño cerebral pequeño indica que los cambios estructurales fueron anteriores al incremento de tamaño (Holloway et al, 2018).
- Hacia los 2 Ma el linaje Homo, cuenta con un tercio inferior del frontal más complejo y moderno en sus circunvoluciones, un volumen craneal superior y asimetrías cerebrales similares a los de los seres humanos actuales.
- Todos los primates antropoides presentan asimetrías morfológicas entre los hemisferios cerebrales.
- En el humano moderno, estas asimetrías se corresponden con especializaciones en el lenguaje simbólico (hemisferio izquierdo) y la integración visuo-espacial (derecho), lo que hace plantear a Holloway la posibilidad de que tales capacidades existiesen ya en los primeros Homo, aun cuando menos desarrolladas. Esta es una idea expresada por primera vez por Tobias.
- La asimetría cerebral tiene también una expresión en términos de dimorfismos sexuales. La relación estrecha madre-hijo en los homínidos, incluso acentuada por una posible prolongación del periodo del embarazo, llevaría a una especialización cerebral relacionada con el corpus callosum. Funcionalmente hablando, las diferencias tenderían a establecer un cerebro masculino especializado en la integración visuo-espacial relacionada con la caza. Por el contrario, las hembras tendrían unas superiores capacidades cognitivas respecto a la comunicación simbólica y la comprensión de la realidad social.
- Estas habilidades serían complementarias más que competitivas.
- La manera como los dimorfismos cognitivos podrían haber afectado a la evolución humana es imposible de plantear como no sea en términos especulativos.
- Aiello y Dunbar (1993) buscan una evidencia medible, como el tamaño cerebral, para rastrear el origen del lenguaje. La conclusión apunta a las primeras fases del género Homo como protagonistas de la aparición del leguaje, con un rápido incremento en la segunda mitad del Pleistoceno Medio, dentro del grado erectus.
- Wynn sostiene, que las capacidades cognitivas dieron un salto brusco con Homo erectus, hasta el punto de atribuir a esa especie una capacidad para manejar datos diversos procedentes del medio, construyendo representaciones complejas del mundo, que sería incluso mayor que la nuestra.
- Los fósiles del grado erectus muestran un amplio rango de variación en su volumen en el espacio y el tiempo. La ausencia de huella del surco fronto-orbital en todos los endomoldes de erectus, junto con la presencia de grandes negativos de Broca indica reorganización de la corteza frontal y evolución de las capacidades lingüísticas. Los endomoldes de erectus también tienen evidencia de una mayor expansión de la corteza prefrontal y de los lóbulos temporales. Queda por aclarar si la expansión siguió una tendencia alométrica general ya presente en los australopitecinos o si podría haber implicado nueva reorganización cortical.
- El sistema frontoparietal es una compleja red cerebral con un papel crucial en la complejidad cognitiva (Jung y Haier, 2007; Caminiti et al, 2015). Los cambios morfológicos son indicativos de reorganizaciones generales del cerebro, incluyendo la conectividad con las áreas subcorticales involucradas en las capacidades lingüísticas (Boeckx y Benítez-Burraco, 2014). Por lo tanto, es interesante que los seres humanos modernos y los neandertales, las dos especies humanas con más rasgos culturales derivados, exhiban características morfológicas específicas de la especie en las áreas frontal y parietal del cerebro (Bruner, 2017).
- Los endomoldes neandertales son similares en tamaño a los de los humanos modernos, pero claramente diferentes en forma. Parece que sus cerebros voluminosos evolucionaron a través de la expansión alométrica de erectus, mientras que los cerebros humanos modernos evolucionaron a lo largo de una nueva vía que implica la expansión de la corteza parietal. Antes del nacimiento, ya se constatan diferencias sustanciales entre la morfología facial y endocraneal del HAM y el neandertal, lo que indica diferencias fundamentales en los programas de desarrollo fetal. Es probable que la divergencia ontogenética prenatal tuviera un efecto decisivo en la organización postnatal del cerebro. Durante la ontogenia temprana postnatal, el cerebro humano moderno adopta una forma esférica, en particular mediante la ampliación de la zona parietal posterior. Más tarde, durante la ontogenia, las trayectorias de desarrollo endocraneal de los humanos modernos y los neandertales son similares, pero más extendidas en este último taxón debido a mayores tasas de crecimiento viscerocraneal. Vanessa R. Paixão-Côrtes et al (2013), han estudiado el genoma completo de HAM, neandertales y denisovanos, para localizar la presencia de genes relacionados con la cognición. El cuadro general obtenido es de similitud de genes cognitivos en humanos extintos y existentes. Según Tristan A. Chaplin et al (2013) la expansión de las areas temporal, parietal (incluyendo el precúneo), corteza prefrontal ventrolateral y corteza cingulada anterior dorsal, todas los cuales son áreas de asociación de alto nivel involucradas en el complejo cognitivo y de comportamiento, sigue un patrón general de escala, presente desde la aparición de los primates.
- Coolidge y Wynn, 2005; Wynn y Coolidge, 2010; Coolidge y Wynn, 2016 proponen que la explosión cultural que caracterizó al sapiens se debió a una mejora en la capacidad de la memoria temporal (Modelo de la Memoria de Trabajo). Los componentes básicos de esta memoria son un centro ejecutivo (corteza prefrontal), un centro de representación visoespacial (lobulillos parietales superiores) y un centro de almacenamiento fonológico (lobulillos parietales inferiores y corteza temporal).
- Baddeley, Gathercole y Papagno (1998) consideran que el almacenamiento fonológico en la memoria de trabajo es un cuello de botella potencial para la adquisición y comprensión del lenguaje. Si el sello distintivo del leguaje moderno es la recursión (Fitch, Hauser y Chomsky, 2005), el almacenamiento fonológico expandido puede haber ayudado al pensamiento recursivo.
- El registro arqueológico sugiere que hace unos 100 ka surgió un importante desarrollo en la cognición humana (aunque el evento genético que mejoró la memoria de trabajo podría haber sido anterior). Klein (2000) y Mithen, 1996 propusieron que hubo una explosión cultural hace unos 50 ka debido a alguna mutación genética.
- La arqueóloga experimental Lyn Wadley (2010) ha estudiado los procedimientos necesarios para fabricar los adhesivos recuperados en herramientas de piedra de la cueva de Sibudu de Sudáfrica (que datan de hace unos 70 ka). La complejidad supera cualquier evidencia anterior y rivaliza con cualquier elaboración de los cazadores recolectores actuales. Además, es claramente una tarea de la función ejecutiva.
 |
| Las teorías antiguas proponían una evolución del cerebro de más simple a más complicado con la sucesiva aparición de nuevas áreas cerebrales. El cerebro de los vertebrados evolucionó a partir de una base común y las distintas áreas se desarrollaron de forma diferencial en las especies según las necesidades específicas. George A. Mashour y Michael T. Alkire (2013). |
Encefalización en Homo.
La espina dorsal rudimentaria de los primeros vertebrados se dividió en cinco partes: medula, cerebro posterior, cerebro medio, diencéfalo (tálamo e hipotálamo) y telencéfalo (hemisferio cerebral). A lo largo de la filogénesis posterior tan solo cambiaron los desarrollos relativos de esas regiones, sobre todo con la expansión de los centros olfatorios del telencéfalo que darán lugar, en los mamiferos, a la aparición del neocórtex.Los mamíferos más primitivos se distinguen de los reptiles por la posesión de un córtex bien desarrollado, con diversas capas de neuronas, en el que Elliot Smith identificó tres regiones, a principios del XX. Dos de ellas podían identificarse con regiones semejantes de los vertebrados inferiores, pero la otra es un rasgo único de los mamíferos, con filogénesis en forma de mosaico: el neocórtex, con cambios evolutivos dispares en los distintos linajes.
Una característica general de todos los primates es el gran desarrollo alcanzado por el neocórtex y dentro de él de las áreas visuales (expansiones extra alométricas, superiores al aumento de tamaño del cuerpo).
Jody A. Creel, 2012, en la evolución de los simios, advierte cambios en la heterocronía y cambios en la citoarquitectura; en la ontogenia, el orangután muestra un patrón más parecido al de los monos que al del chimpancé y humano.
Según Dante Mantini et al (2013), dos redes situadas en las regiones corticales frontoparietales son nuevas en los humanos evolutivamente hablando. Se supone que están relacionadas con aspectos específicos de nuestra inteligencia.
Según Dwight Read (2014) en el sapiens la memoria de trabajo es de mayor tamaño que en el chimpancé, lo que da lugar también a diferencias cualitativas, como la emergencia de la recursividad.
Mary-Claire King y Allan Wilson desarrollaron la hipótesis del regulador de llamada según la cual son las diferencias en la actividad de los genes - la velocidad a la que se desencadena la formación de ARN y proteínas por los genes - las responsables de las diferencias en la morfología cerebral, el comportamiento y la cognición. En comparación con los chimpancés, las diferencias cuantitativas en ambos ARN y la producción de proteínas es hasta seis veces más alta en los seres humanos. De ello se deduce que la actividad de los genes del cerebro humano ha aumentado enormemente durante su evolución. Ignoramos cómo y cuándo se produjo este cambio.
Randy L. Buckner y Fenna M. Krienen (2013) han estudiado la emergencia de la mente humana. Según su hipótesis de la atadura (tether hypothesis) el cambio se produjo cuando, debido al crecimiento del cerebro, las conexiones entre las neuronas se rompieron o desataron al alejarse entre sí, de forma que pudieron formar nuevos circuitos y originar nuevas cortezas de asociación.
La tendencia hacia la encefalización comenzó con la remodelación biomecánica que se puede observar ya en Ardipithecus. Australopithecus africanus disponía de una capacidad encefálica en torno a los 450 cc. Homo habilis, con un tamaño corporal similar al A. Africanus , 600. El cráneo KNM-ER 1470 tiene una capacidad de 752 cc., está datado hace 1,9 Ma y atribuido a Homo rudolfensis. Para José María Bermúdez de Castro, (La evolución del talento) el incremento en el coeficiente de encefalización pudo producirse:
- Gracias a un aumento en la en la tasa de crecimiento del cerebro durante la gestación. La pelvis del Homo habilis era relativamente más ancha que la del sapiens, y el aumento del tamaño del cerebro no debió suponer problema alguno en el parto.
- El problema obstétrico consiste en explicar un aumento del volumen craneal al mismo tiempo que aumentaban los problemas del parto debido a las implicaciones de la bipedación. Parece que lo adaptativo hubiese sido lo contrario. Del estudio del cráneo del Niño de Taung, Dean Falk et al (2012) deducen que la fontanela anterior se soldaba tardíamente, tal como sucede en los humanos y esta adaptación, que facilita el parto, pudo permitir también un aumento del volumen del cerebro.
- La morfología de los huesos de la pelvis, el insquion y el pubis, indican que en el Australopithecus afarensis el parto era parecido al de las humanas modernas, con rotación de la cabeza del feto y una trayectoria curva del canal del parto. El feto tenía un 50% de posibilidades de nacer con la coronilla orientada hacia su madre, lo que dificulta el parto en solitario.
- Los problemas crecieron cuando los adultos alcanzaron un tamaño cerebral de 800 cc, con Homo erectus. Debido a la morfología del útero, la vagina y la pelvis, el parto tuvo que producirse casi siempre con la coronilla del feto orientada hacia la madre. El canal del parto tiene un diámetro máximo de 13 cm. La asistencia al parto es casi obligada. A partir de entonces los incrementos en el tamaño del cerebro tuvieron que producirse fuera del útero de la madre y esto pudo llevar a un aumento de la altricialidad o inmadurez de la cría en el momento del parto. La relación entre las masas del encéfalo en el adulto y neonato es de 3,7 en humanos mientras que en el chimpancé es de 2,8.
- Gracias al aumento en la duración de la infancia.
En los simios antropomorfos, los lóbulos frontales se sitúan detrás de los ojos. En los humanos más arcaicos, se encuentran en una posición intermedia. Solo en HAM y neandertales los lóbulos frontales están directamente apoyados en las órbitas. En relación a los neandertales y los primeros sapiens, los HAM tenemos lóbulos parietales más largos y abultados que corresponden con la posición del precúneo.
Los neandertales tenían las superficies parietales dorsales más anchas y abultadas lateralmente que los homininos arcaicos. Justo debajo de esta superficie, se encuentra el surco intraparietal.
En el HAM observamos un aumento en la complejidad de la red vascular, seguramente relacionada con la regulación térmica. Afecta sobre todo a la superficie parietal.
Mecanismos genéticos
Mecanismos genéticos implicados (Rosales-Reynoso et al, 2018):- Selección positiva en regiones genéticas codificantes de proteínas que conducen a cambios en la secuencia y actividad de proteínas existentes.
- Cuando se comparan los humanos y los chimpancés, la mayoría de las proteínas presentan al menos una diferencia en sus aminoácidos. Algunas de estas diferencias han tenido efectos funcionales importantes que han contribuido a ciertas características cerebrales humanas específicas. Cuando la selección positiva actúa sobre las mutaciones en regiones codificadoras que alteran la secuencia de la proteína, frecuentemente deja huella en los genes afectados. El principal enfoque en el estudio de la genética y sobre todo en la evolución del cerebro humano ha sido la identificación de genes que han mostrado selección positiva en comparación con el resto de primates.
- La microcefalia primaria se manifiesta como una reducción en el tamaño del cerebro sin otras anormalidades en la arquitectura o en la formación del giro. Típicamente, la reducción en el tamaño de la corteza cerebral es mucho más severa que la de otras regiones del cerebro. Por lo tanto, la microcefalia primaria puede ser considerada como una condición atávica que recapitula algunos aspectos de las etapas evolutivas anteriores. Se han identificado seis regiones del genoma humano (MCPH1 a MCPH6); además se han asociado mutaciones nulas en 4 loci: microcefalina (MCPH1); proteína 2 reguladora asociada a la subunidad regulatoria CDK5 (CDK5RAP2); microcefalia asociada al uso mitótico anormal (ASPM) y proteína centromérica (CENPJ). El gen ASPM ha sufrido selección positiva durante todo el linaje de los primates a los seres humanos e incluye tanto el linaje de la separación de los grandes simios del Viejo Mundo hasta el linaje dentro de los grandes simios, el cual nos incluye como seres humanos. Por otra parte, el gen de la microcefalina se ha caracterizado por una marcada selección positiva principalmente en el linaje de los primates ancestrales hasta los grandes simios Así mismo los genes CDK5RAP2 y CENPJ han mostrado mayores tasas de sustituciones no sinónimas en primates comparadas con los roedores y CDK5RAP2 mostró específicamente altas tasas de cambios en los linajes del chimpancé y los humanos.
- Otro ejemplo relacionado con la proliferación de precursores neuronales es el gen polipéptido 1 de activación de adenilato ciclasa (ADCYAP1), que también ha sufrido una acelerada evolución en su secuencia proteica en los seres humanos. ADCYAP1 regula la transición de la proliferación de estados diferenciados durante la neurogénesis.
- Un segundo aspecto del desarrollo neuronal que podría haber sido sustrato para la selección positiva en los seres humanos es el direccionamiento del axón. El gen auxiliar 1 del sitio de integración Abelson (AHI1) es uno de los genes asociados a una rara malformación cerebral denominada síndrome de Joubert; el gen AHI1 participa direccionando los axones desde el cerebro hasta la médula espinal. En ese sentido, tanto los genes ASPM, ADCYAP1 y AHI1 muestran una tasa acelerada de cambios no sinónimos en la secuencia a lo largo del linaje humano a partir de la divergencia de los chimpancés.
- Otro ejemplo es el gen Sonic Hedgehog (SHH), el cual es altamente conservado durante el desarrollo. Este gen codifica para una molécula de señalización que tiene un papel central durante el desarrollo de muchos tejidos, especialmente en el sistema nervioso y esquelético. La proteína SHH está constituida por 2 dominios funcionales: el péptido señal y una región autocatalítica; ambos dominios se encargan de cortar el péptido señal. El dominio autocatalítico ha mostrado una marcada tasa de evolución en su secuencia proteica en primates comparada con otros mamíferos, y más aún en la secuencia proteica de los humanos, donde los principales cambios involucran ganancia de ciertos aminoácidos como serinas y treoninas, los cuales son sustratos potenciales para modificaciones postraduccionales, por lo que este gen SHH se ha propuesto como el principal autor de la evolución del sistema nervioso y/o sistema esquelético desde los primates hasta el humano.
- Finalmente, otro gen relacionado en estos procesos es monoamino oxidasa A (MAOA), el cual codifica para una enzima mitocondrial que cataboliza varios neurotransmisores, entre los que se incluyen la dopamina, la serotonina y la norepinefrina. Alteraciones funcionales en este gen pueden tener potencialmente numerosas consecuencias fisiológicas y conductuales. Se sugiere que mutaciones no sinónimas en este gen provocaron un cambio funcional en la enzima, dando lugar a la divergencia humano-chimpancé.
- RNF213 muestra una evolución acelerada en humanos, chimpancés y gorilas. Pudo facilitar un flujo sanguíneo adicional para permitir grandes cerebros (Scally et al, 2012).
- Duplicación de genes previamente existentes.
- La generación de nuevos genes en el genoma es desencadenada por eventos de duplicación, a menudo dentro de grandes familias de genes que quizás tienen predisposición a tales eventos. La relajación de la restricción evolutiva en los genes duplicados proporciona una oportunidad única para neofuncionalización o subfuncionalización. La «neofuncionalización» involucra la adquisición de nuevas características por uno o ambos genes duplicados, mientras que la «subfuncionalización» implica la repartición de la función ancestral entre los genes duplicados. Los recientes avances en la secuenciación completa de genomas de múltiples especies de primates, acopladas con nuevas tecnologías como la hibridación genómica comparativa, ha facilitado la identificación de genes duplicados durante la evolución humana
- La primera familia de genes para los cuales existe evidencia sustentable de duplicación génica, seguida por neofuncionalización, es la familia «Morfeo». Esta familia de genes es tan amplia en los linajes de humanos y los grandes simios que ha sido acompañada por una intensa selección positiva en las secuencias de proteínas codificadas. La función de los genes Morfeo es aún desconocida y no está claro si la adaptación drástica de esta familia de genes a través de la evolución tiene que ver con el proceso de encefalización humana.
- Según Franck Polleux y Evan Eichler (2012), unos 30 genes se duplicaron de forma selectiva en los seres humanos. SRGAP2 se duplicó al menos dos veces: hace 3.5 Ma y hace 2,5 Ma, esta vez de forma parcial, dando lugar a una innovación genética que podría haber ayudado a poner en marcha los cambios evolutivos en el desarrollo del cerebro y coincide en el momento de la transición entre Australopithecus y Homo. Esta innovación habría acelerado la migración neuronal dando más tiempo para la creación de conexiones neuronales. SRGAP2C es responsable de un aumento en el número de conexiones entre las neuronas (Suzuki et al, 2018).
- Tal cambio trascendental ocurrió al mismo tiempo que la inhibición del gen MYH16, de forma que los músculos relacionados con la masticación disminuyeron su tamaño y dejaron de limitar el crecimiento cerebral (Stedman et al, 2004; Rosales-Reynoso, 2018).
- Según Laura J. Dumas et al (2012) hay una correlación entre el número de copias del dominio de la proteína DUF1220, en la familia de genes NBPF, y el tamaño del cerebro en humanos (patologías) y simios. Los humanos disponemos por lo general de 270 copias, los chimpancés 125, los gorilas 99, los titíes 30 y los ratones 1. Por ello este gen es candidato a efector de la evolución biológica y la variación patológica.
 |
| Dendrograma de grandes simios en relación con NOTCH2NL. Fiddes et al (2018) |
- Fiddes et al (2018) descubrieron que el gen NOTCH2NL (genes similares a N terminal de homólogos de muesca 2 “Notch Homolog 2 Nterminal-Like), presente en los humanos actuales, falta en el macaco. Al reconstruir la historia evolutiva de este gen, llegaron a la conclusión de que hace unos 14 Ma, parte de un gen ancestral NOTCH2 se copió por error, con duplicación parcial, reparación y conversión. El nuevo "gen" era incompleto y no funcional, pero hace ca 3 Ma, poco antes de que los cerebros de los ancestros humanos comenzaran a expandirse, se insertó una pieza adicional de NOTCH2 en esta copia, haciendo que el gen fuera funcional. Posteriormente, ese gen NOTCH2NL activo se duplicó dos veces más, produciendo tres genes activos de NOTCH2NL en un extremo del cromosoma 1 humano y una copia inactiva en el otro extremo. Suzuki et al (2018) y Florio et al (2018) descubrieron que este gen retrasa la especialización de las células madre de la corteza cerebral responsables de la producción de neuronas. Este retraso produce muchas más copias de células madre y en consecuencia se obtienen más neuronas.
- Los tres genes NOTCH2NL activos (NOTCH2NLA, NOTCH2NLB y NOTCH2NLC) se ubican en una región inestable, en el medio del ADN implicado en el autismo, la esquizofrenia y el síndrome de retraso del desarrollo. La sobreexpresión de NOTCH2NL retrasa la diferenciación neuronal, mientras que su eliminación lo acelera. Las duplicaciones, reparaciones y conversiones de uno o ambos genes NOTCH2NLA y NOTCH2NLB pueden estar relacionadas con macrocefalia, mientras que las deleciones de uno o ambos se han asociado a microcefalia.
- Otra familia de genes denominados MRG, los cuales codifican para un grupo de receptores acoplados de proteínas G, se expresan significativamente en neuronas nociceptivas de la médula espinal y son implicadas en la modulación de la nocipercepción. La presencia de múltiples copias de los genes MRG en humanos, probablemente es el resultado de la amplificación del gen después de la divergencia humano-ratón, ya que las secuencias de las copias en humanos revelan fuerte evidencia de selección positiva en las regiones de los genes que codifican para los dominios de unión al ligando extracelular.
- Otro ejemplo en el que la consecuencia funcional de un evento de duplicación es evidente, es el nacimiento del gen glutamato deshidrogenasa 2 (GLUD2). Este gen surgió de la retrotransposición o de la reintegración del ARNm de solo un precursor ancestral: GLUD1. En la mayoría de las especies de mamíferos, GLUD1 es el único gen que codifica para la proteína glutamato deshidrogenasa, la cual en el cerebro cataliza el reciclaje del neurotransmisor excitatorio glutamato. El evento de retrotranscripción ocurrió en el linaje de los simios, después de su divergencia de los monos del Viejo Mundo, dando lugar al gen GLUD2, el cual codifica para otro gen glutamato deshidrogenasa específico en los simios y los humanos. Aunque el gen GLUD1 es ampliamente expresado en muchos tejidos, la expresión de GLUD2 es específica del tejido nervioso y testicular.
- Deleción genética.
- La pérdida de genes en un organismo puede tener consecuencias drásticas o deletéreas, por lo que este es un evento mucho más raro durante la evolución. En algunos casos, sin embargo, la pérdida de genes puede ocurrir debido a cambios selectivos con el tiempo.
- El mejor ejemplo conocido de pérdida de genes durante la evolución humana es la familia de genes de los receptores olfatorios (RO). Se ha estimado que el ratón tiene aproximadamente 1.200 genes con funciones de RO, en tanto que en el humano solo existen cerca de 350 de estos genes. Esta cantidad reducida de genes es debida principalmente a los seudogenes (reliquias no funcionales de los genes ancestrales). Esta degeneración en los genes RO no es única en humanos y parece afectar también varias especies de primates. Recientemente se ha demostrado una selección positiva en algunos genes RO en el linaje humano.
- Otro ejemplo interesante de pérdida de genes que podría haber desempeñado un papel importante en la evolución del cerebro humano es el gen cadena pesada 16 de la miosina (MYH16); este gen codifica para la cadena pesada de la miosina, proteína presente en el músculo esquelético. En primates no humanos, MYH16 se expresa exclusivamente en músculos de la cabeza, incluidos los músculos encargados de la masticación; en humanos, una mutación en este gen produce desplazamiento del marco de lectura, lo que resulta en la pérdida de su función. En ese sentido, al realizarse comparaciones con otros primates, los humanos tienen un sistema para la masticación poco desarrollado, sugiriendo que la pérdida del gen MYH16 fue parcialmente responsable de ello.
- Cambios en la expresión génica a través de modificaciones en las secuencias reguladoras de diferentes genes.
- Se ha postulado que cambios en la expresión génica hayan desempeñado un papel importante en la aparición del fenotipo humano. En particular, ha sido argumentado que pequeños cambios en elementos regulatorios no codificantes pudieran fuertemente impactar en los patrones de expresión espacial y temporal de genes del desarrollo, lo que podría tener profundos efectos fenotípicos.
- Una estrategia utilizada para establecer cómo los cambios en la expresión génica han contribuido a la evolución humana es comparar regiones regulatorias cis de genes relacionados con el cerebro, para identificar aquellos genes que han sufrido selección positiva. El gen prodinorfina (PDYN) codifica para un precursor de un neuropéptido opioide involucrado en muchos procesos neurales. Un elemento cis regulatorio ubicado corriente arriba de este gen exhibe una tasa rápida de cambios en su secuencia en el linaje humano después de la divergencia de los chimpancés, lo que demuestra los efectos de la selección positiva. Además de secuencias reguladoras cis, los cambios en las secuencias proteicas de factores de transcripción pueden tener profundos efectos en la expresión de los genes que regulan. Varios estudios han encontrado un aumento significativo de factores de transcripción entre los genes que han experimentado selección positiva en sus regiones codificantes para las proteínas, aunque no es claro si los patrones de expresión de sus genes diana se ven afectados.
- Cambios en la síntesis de ARNs no codificantes.
- Durante una exploración a gran escala de elementos cis-reguladores, se encontró el primer ARN no codificante humano seleccionado positivamente (Pollard et al., 2006). El gen para este ARN no codificante fue el llamado región humana acelerada 1 (HAR1), el cual es expresado en las neuronas del desarrollo del neocórtex humano. El análisis evolutivo realizado reveló que, a pesar de que este gen mide solo 118 pb, contiene 18 cambios en el linaje humano desde la divergencia de los chimpancés.
- J. Lomax Boyd et al (2015) han obtenido las principales diferencias en potenciadores genéticos entre los chimpancés y los seres humanos y luego han visualizado sus respectivas contribuciones al desarrollo temprano del cerebro, implantando en embriones transgénicos de ratón las correspondientes variantes. Los potenciadores candidatos fueron denominados HARE (human-accelerated regulatory enhancer) de 1 a 6. Según los hallazgos, cuando se introduce en un embrión de ratón el potenciador humano HARE5, da lugar a un cerebro un 12% más grande que cuando se introduce la secuencia HARE5 de los chimpancés, siendo el neocórtex la región cuyo volumen más aumenta. Los HARE5 humano y del chimpancé se difierencian en sólo 16 letras en su código genético. Sin embargo, en embriones de ratón, los autores encontraron que el potenciador humano se activaba antes en el desarrollo y era más activo en general que el promotor del chimpancé. HARE5 potencia el gen FZD8, un receptor en la vía de señalización de Wnt del grupo Frizzled (fz) que dirige la polaridad celular, desarrollo embrionario y muchos otros procesos en organismos adultos y en desarrollo.
Modificaciones epigenéticas
Cada individuo logra su «individualización» a través de sus interacciones con el entorno (Rosales-Reynoso et al, 2018). Se cree que en los seres humanos estos mecanismos epigenéticos son los responsables de esos 900 g de materia gris que tenemos en exceso en nuestro cerebro y de la forma en que las áreas cerebrales se distribuyen. Sin entrar en mayor detalle respecto al genoma humano, debemos recordar que contiene entre 20 y 25 mil genes, los cuales codifican para el conjunto de proteínas que forman nuestra estructura bioquímica. Las secuencias codificadoras de proteínas representan menos del 2% de nuestro genoma; el otro 98% del genoma humano está constituido principalmente por secuencias que regulan la expresión génica, incluyendo secuencias de ARN no codificantes con funciones regulatorias.Como se mencionó previamente, los seres humanos (H. sapiens) y los chimpancés (P. troglodytes) nos diferenciamos en tan solo un 1,23% de la secuencia genómica. Además de los cambios de estructura y función de las proteínas, las diferencias en los patrones de expresión génica son considerados fundamentales para la diversificación fenotípica. Estudios comparativos han identificado numerosas modificaciones epigenéticas en regiones reguladoras. La metilación de citosinas en sitios CpG es la principal modificación epigenética en el ADN genómico, y es capaz de regular la expresión génica de numerosas secuencias codificadoras. En mamíferos, los sitios CpG que contienen citosinas metiladas se caracterizan por una elevada tasa de mutabilidad. Las islas CpG son regiones de 0,5-2 kilobases que albergan muchos sitios CpG, generalmente localizados en regiones promotoras no metiladas de los genes. La metilación de estas regiones es un proceso heredable a través de las divisiones celulares, lo que es importante para el mantenimiento de los patrones de expresión. Se observan niveles de metilación variables en regiones adyacentes a las islas CpG durante el desarrollo embrionario, la carcinogénesis y la reprogramación celular. En humanos y chimpancés, el 12-18% de los genes que muestran diferencias de expresión tienen también diferencias de metilación.
- Zeng et al. (2012) identificaron 1055 genes en el córtex humano que están significativamente menos metilados que en la corteza del chimpancé. Muchos de estos genes hipometilados humanos mostraron una mayor expresión en comparación con sus homólogos de chimpancé. Además, este conjunto incluye una serie de genes asociados con el autismo, como GABRA2, GABRG1 (genes del receptor GABA) y SLC6A4 (el gen transportador de la serotonina). En los seres humanos, la influencia de un gen en la cognición social puede estar implícita cuando la disfunción genética se asocia con trastornos de la comunicación o rasgos del espectro autista, como la falta de empatía o habilidades sociales (Ragsdale y Foley, 2018).
 |
| Mediante la comparación de las áreas homólogas cerebrales de especies y ancestros de mamíferos y primates, puede construirse un árbol filogenético. Las relaciones espaciales generales entre áreas sensoriales primarias son consistentes a través de especies de mamíferos y la capa cortical del primate está ocupada principalmente por zonas que se encuentren entre las áreas sensoriales primarias y secundarias. Randy L. Buckner y Fenna M. Krienen (2013) |
Ventajas de la encefalización
Nuestro encéfalo representa un 2,5% de la masa corporal, pero su consumo asciende a un 22% de la tasa metabólica basal (1,33 kg pesa nuestro cerebro y su consumo energético equivale al de 25 kg de músculo esquelético en reposo). La encefalización tiene un coste metabólico y por tanto debemos preguntarnos cuáles son los superiores beneficios en términos adaptativos de un mayor tamaño del encéfalo.- Las especies más encefalizadas, pueden explotar territorios más amplios y con ello alimentarse de recursos que requieren de una mayor capacidad de predicción.
- Hipótesis del forrajeo (extractive foraging hypothesis)
- Katherine Milton y Katherine Gibson entre otros han atribuido las capacidades cognitivas mejoradas de los primates a las ventajas de poseer un mapa cognitivo espacio-temporal para el forrajeo debido a que los alimentos se distribuyen por el espacio y las estaciones.
- Katherine Gibson considera además que la obtención de alimentos que se ocultan de la vista, a menudo difíciles de extraer, como nueces, raíces y tubérculos subterráneos, y que requieren un delicada manipulación para recuperar con éxito, pueden haber jugado un papel en el aumento de la capacidad cognitiva de los primates.
- La relación depredador - presa conduce a una carrera armamentística, en la que el aumento de tamaño del encéfalo provee de más y mejores estrategias de caza o huída.
- El mayor tamaño del encéfalo permite dedicar una parte a las habilidades sociales. Vivir en grupo disminuye los factores de riesgo, pero exige comportamientos más complejos (teoría del cerebro social).
Cambios en la organización del cerebro. Lenguaje.
El hecho de la encefalización creciente no es el único fenómeno
notorio que pueda detectarse en la filogénesis de los homínidos. Además del aumento
de tamaño relativo del ceerebro, que es una cuestión en esencia cuantitativa, se producen también
cambios en la organización que mantuvieron e incrementaron su eficiencia. Para JB Smaers y C. Soligo (2013), la reorganización del cerebro fue la principal fuerza evolutiva y la corteza prefrontal juega el papel más importante en la explicación de los cambios evolutivos de los antropoides. Por otra parte, el tamaño del cerebro y la organización cortical muestran un alto componente hereditario en los chimpancés, mientras que en los humanos aumenta la plasticidad y la influencia del medio ambiente en el desarrollo neurológico (Aida Gómez-Robles et al, 2015).
- Semendeferi y Hanna Damasio (2000) compararon en distintos hominoideos los volúmenes relativos de tres áreas cerebrales, la frontal, la temporal y la parieto-occipital (definidas por el sulcus central y la fisura de Silvio) para concluir que los lóbulos temporales, reconocidos por su importancia para la audición, visión y muchos otros procesos cognitivos superiores como la memoria y el lenguaje, aparecen anormalmente grandes en nuestra especie, lo que hace pensar que existe una presión selectiva en el género Homo para el desarrollo de esta parte del cerebro.
- Robert A. Barton y Chris Venditti (2013), han confirmado la expansión alométrica de los lóbulos frontales.
- El estudio de Rilling e Insel (1999) indica que la girificación es, en términos generales, alométrica en los hominoideos: cuanto más grande es el cerebro, más girificado está el córtex. Pero en las secciones longitudinales horizontales Rilling e Insel detectaron una girificación extraalométrica en la “rebanada” que corresponde en gran parte al córtex frontal y prefrontal en Homo sapiens. Eso indica que el desarrollo último del lenguaje, la capacidad estética y los juicios morales puede tener que ver con esas áreas. Zeki (1999) sugiere que el arte pictórico podría deberse a ciertos cambios producidos en las mismas áreas visuales, es decir, en la zona occipital del cerebro.
- Schoenemann, Sheehan y Glotzer (2005) encontraron mayor materia blanca prefrontal en los humanos y Bufill y Carbonell (2004) una mayor y tardía mielinización. Esto indica una mayor interconexión.
- La diferenciación funcional (A. Rivera; pub on-line, 2014) conlleva a que nuestra especie tendría una mayor capacidad funcional de dos tipos (fundamentales en la conducta humana) (Ardilla y Ostrosky-Solis, 2008):
- Metacognitivos (área dorsolateral de la corteza prefrontal), para procesar la información, asimilarla y utilizarla para mejorar su conducta, mediante el mayor desarrollo de sus funciones ejecutivas, imprescindibles para la organización de todo tipo de conducta y lenguaje, y al aumento de las capacidades de abstracción y simbolismo.
- Emocionales (área ventromedial de la corteza prefrontal), que coordina la cognición y la emoción. En ese sentido, la función principal del lóbulo prefrontal es encontrar justificaciones aparentemente aceptables para los impulsos límbicos (los cuales constituyen las “funciones ejecutivas emocionales”).
- El estudio comparativo de Falk et al (2000) indica una medida relativa igual para el lóbulo central en humanos actuales, gorilas y chimpancés, pero con diferencias significativas a favor de los humanos en algunas subregiones del lóbulo central. Esto quiere decir que el lóbulo frontal humano se ha reorganizado en comparación con el de los grandes simios africanos. Los lóbulos frontales de los parántropos muestran una forma tipo-simio mientras que la expansión frontal y temporal de Australopithecus africanus se acerca más a Homo.
- La primera, la aparición en Homo habilis de una organización neurológica “esencialmente humana”.
- La segunda, un continuo y rápido aumento del índice de encefalización dentro del género Homo.
- Reducción relativa de la corteza visual primaria y el cambio asociado del surco semilunar hacia los polos occipitales.
- Ampliación mediolateral de los lóbulos frontales.
- Expansión de los polos temporales, asociada con un mayor nivel de abstracción, razonamiento matemático y económico más complejo y mayor autoconciencia.
- Desaparición del surco fronto-orbital simiesco de la superficie cortical externa y la superficie endocraneana asociada.
 |
| Globularización del cerebro. Bruner e Iriki (2015). |
Bruner y Lozano (2014), Bruner e Iriki (2015), Bruner et al (2016) y Bruner, Lozano y Lorenzo (2016) Bruner et al (2018) recuerdan que las evidencias paleoneurológicas sugieren que las áreas del cerebro que más han cambiado a lo largo de la evolución humana son las relacionadas con la integración visual-espacial (surco intraparietal y precúneo):
- Los neandertales muestran un abultamiento lateral de las áreas parietales superiores que no aparece en erectus ni heidelbergensis. Para los autores puede haber una relación entre estos cambios morfológicos y la utilización de las técnicas líticas Levallois. El uso de la boca como una tercera mano pudo haber sido el resultado de una organización cerebral insuficientemente ajustada a la complejidad cultural, lo que requirió el reclutamiento de elementos corporales adicionales.
- La forma globular del cerebro del HAM es debida a una patente dilatación espacial de toda la superficie parietal en una fase post-natal muy temprana, lo que no sucede en los monos ni en los neandertales.
Tres regiones de la corteza y las estructuras endocraneales relacionados son de especial interés para la paleoneurología: La corteza prefrontal, las áreas de Broca y Wernicke y los lóbulos temporales y los lóbulos occipitales.
- La corteza prefrontal, que se considera generalmente como una región clave para el conocimiento avanzado.
- Las áreas de Broca y de Wernicke y los lóbulos temporales, que están implicadas en el procesamiento del lenguaje (Flinker et al, 2015). El paso de hominoideos primitivos a seres del género Homo tiene que ver con el desarrollo de habilidades de modificación del entorno: construcción de herramientas, dominio del fuego, cooperación y división del trabajo etc. Pero estas actividades necesitan de un lenguaje con el que la transmisión de experiencias se haga al margen de la herencia genética a través de una memética cultural. Los sapiens cuentan con un desarrollo muy acusado de las áreas temporales y prefrontales relacionadas con el procesamiento de la comunicación verbal y las tareas semánticas. Cabría sustentar en el desarrollo de tales áreas las especulaciones acerca del origen del lenguaje. Tobias ha venido defendiendo una expansión temprana de estas áreas. Comparando los endocráneos de Homo habilis con los de Australopithecus africanus, Tobias detectó no solo un desarrollo de las áreas de Broca y Wernicke sino también del lóbulo frontal, que anticiparía el incremento de volumen de esa zona en los homínidos de grado erectus.
- Para Holloway (1983), la paleoneurología y el registro fósil no pueden dar ninguna solución al debate sobre la evolución del lenguaje (Bruner, 2017).
- En los cerebros humanos modernos, el área de Broca a menudo tiene un negativo en el endocráneo. Su presencia en endomoldes homínidos fósiles se ha interpretado como evidencia de la evolución temprana del lenguaje articulado. Los estudios basados en MRI, sin embargo, han demostrado que los cerebros de los chimpancés también muestran un área de Broca, aunque su misión funcional no sea la misma, ya que la citoarquitectura de sus áreas y bordes son algo diferentes de las de los seres humanos.
- Más allá de los indicios extraídos de los moldes endocraneales, apenas puede decirse nada que no sea en términos especulativos. Lieberman sostiene que el habilis tendría, en el mejor de los casos, un lenguaje no totalmente moderno mientras que Tobias cree en una creciente complejidad en los aspectos conceptual y sintáctico, junto con una ampliación en el tiempo del repertorio de fonemas. Todas estas etapas entrarían dentro de lo que cabe considerar como “lengua humana hablada”.
- El aprendizaje del lenguaje está correlacionado con la mielinización del fascículo arqueado, que une el área de Wernicke y el área de Broca.
- Será necesario disponer de estudios más detallados para localizar de forma precisa las funciones cerebrales que intervienen en los procesos cognitivos relacionados con el habla antes de poder decir la última palabra. Así por ejemplo, Iain DeWitt y Josef P. Rauschecker (2011) han comunicado que tras analizar 115 estudios de resonancia magnética funcional y tomografía por emisión de positrones, el procesamiento del habla se produce en una nueva área unos 3 cm más cerca de la parte frontal del cerebro que el área de Wernicke, al otro lado de la corteza auditiva. Esta nueva ubicación parece concordar con la recientemente descubierta área en primates no humanos, lo que sugiere que el origen del lenguaje entre primates y humanos es más cercano de lo que se creía. Por otra parte, el lenguaje implica una amplia gama de regiones corticales adicionales, lo que indica la naturaleza altamente interconectada de la funciones cerebrales cognitivamente exigentes. Estos estudios muestran que el lenguaje no es sólo una cuestión de módulos corticales especializados, sino de cómo se interconectan los módulos entre sí. De especial relevancia es la obra de Stout y Chaminade (2012), que puso de manifiesto que una parte importante de los circuitos neuronales implicados en la producción del lenguaje también se activan durante la producción de herramientas de piedra. El análogo en el chimpancé del área de Broca está implicado en los gestos, lo que presta apoyo a la hipótesis de que la evolución del lenguaje se correlacionó con la evolución de las habilidad habilidades manuales, en especial la producción de herramientas de piedra.
- Por otra parte, se ha discutido mucho acerca de la influencia de factores innatos en el procesamiento del lenguaje. Mahdi Mahmoudzadeh et al (2013), han estudiado las respuestas cerebrales al habla en fetos de 28 a 32 semanas de gestación, cuando aún no se ha completado la organización cortical en capas, observando varios puntos de semejanza con las redes neuronales lingüísticas de los adultos, lo que confirma la existencia de dichos factores innatos. La hipótesis sound symbolism bootstrapping propone que existe una relación entre el sonido de una palabra y su significado y que los infantes preverbales cuentan con una capacidad sinestésica en forma de cableado cerebral transversal (integración intermodal) que les ayuda a asociar el sonido con el referente (Imai y Kita, 2014; Bankieris y Simmer, 2015).
- Los lóbulos occipitales, que comprenden las principales áreas de procesamiento de la información visual.
- En los grandes simios, el surco semilunar representa la delimitación anterior de la corteza visual primaria. Esta delimitación tiene su negativo como una impresión superficial en algunos endomoldes.
- En los cerebros humanos modernos, el surco semilunar se encuentra más posteromedialmente, lo que indica un importante desplazamiento de las tareas de procesamiento de información visual a las áreas corticales. En los seres humanos modernos, el surco semilunar es poco visible, muy variable, y sólo parcialmente representado en la superficie cortical externa. Además, no representa el borde anterior de la corteza visual primaria.
Lateralización
El patrón primate señala que a un cerebro más grande le corresponde mayor asimietría. Sin embargo, el estudio de las asimetrías estructurales cerebrales como sustratos anatómicos de las asimetrías funcionales en seres humanos existentes, grandes simios y homínidos fósiles es de gran importancia en la comprensión de las bases estructurales de la cognición humana moderna.
El hemisferio derecho es ligeramente mayor que el izquierdo, extendiéndose más hacia el polo frontal, mientras que el izquierdo se extiende más hacia el polo occipital (patrón petalial). Los lóbulos frontal y parietal derechos son a menudo más gruesos que los izquierdos, mientras que el occipital y el área interna del lóbulo temporal izquierdos, así como la mitad izquierda de la parte lateral de la corteza prefrontal son más grandes que los derechos. El planum temporal del lóbulo temporal, parte del área de Wernicke, es mayor en el lado derecho en la mayoría de los humanos. La pendiente de la Cisura de Silvio, que define el borde superior del lóbulo temporal, es más pronunciada en el lado derecho. El el surco temporal superior es más profundo en el hemisferio derecho, en la zona ventral del giro de Heschl; esta asimetría es una apomorfia humana (François Leroy et al, 2014).
La lateralidad y la asimetría del cerebro son consideradas como exclusivas de los humanos, y se asocian con una especialización del hemisferio izquierdo para el lenguaje (incluido el de gestos), la lógica y pensamiento, los gestos y el uso de herramientas ( los humanos actuales muestran una amplia preferencia por la mano derecha, 85-90%; Chui y Damasio, 1980, Annett, 1985, Amunts et al., 2000,Fagard, 2004, Raymond y Pontier, 2004, Cashmore et al., 2008, Uomini, 2009 y Papadatou-Pastou, 2011) y del hemisferio derecho para la creatividad, la intuición, la visión y la atención. Sin embargo, las asimetrías están muy extendidas entre los animales, lo que apoya la teoría de la evolución gradual de funciones como el lenguaje y el uso de herramientas (Forrester et al, 2015).
El hemisferio derecho es ligeramente mayor que el izquierdo, extendiéndose más hacia el polo frontal, mientras que el izquierdo se extiende más hacia el polo occipital (patrón petalial). Los lóbulos frontal y parietal derechos son a menudo más gruesos que los izquierdos, mientras que el occipital y el área interna del lóbulo temporal izquierdos, así como la mitad izquierda de la parte lateral de la corteza prefrontal son más grandes que los derechos. El planum temporal del lóbulo temporal, parte del área de Wernicke, es mayor en el lado derecho en la mayoría de los humanos. La pendiente de la Cisura de Silvio, que define el borde superior del lóbulo temporal, es más pronunciada en el lado derecho. El el surco temporal superior es más profundo en el hemisferio derecho, en la zona ventral del giro de Heschl; esta asimetría es una apomorfia humana (François Leroy et al, 2014).
La lateralidad y la asimetría del cerebro son consideradas como exclusivas de los humanos, y se asocian con una especialización del hemisferio izquierdo para el lenguaje (incluido el de gestos), la lógica y pensamiento, los gestos y el uso de herramientas ( los humanos actuales muestran una amplia preferencia por la mano derecha, 85-90%; Chui y Damasio, 1980, Annett, 1985, Amunts et al., 2000,Fagard, 2004, Raymond y Pontier, 2004, Cashmore et al., 2008, Uomini, 2009 y Papadatou-Pastou, 2011) y del hemisferio derecho para la creatividad, la intuición, la visión y la atención. Sin embargo, las asimetrías están muy extendidas entre los animales, lo que apoya la teoría de la evolución gradual de funciones como el lenguaje y el uso de herramientas (Forrester et al, 2015).
- La dominancia del hemisferio derecho para la emoción parece estar presente en todos los primates hasta ahora investigados, lo que sugiere una continuidad evolutiva que se remonta por lo menos a 30-40 Ma.
- El predominio del hemisferio izquierdo para la vocalización se ha demostrado en ratones y ranas. Alrededor de dos tercios de los chimpancés son diestros, especialmente en gesticular y lanzar. El hemisferio izquierdo de los chimpancés muestra dos áreas corticales homólogas a las principales áreas del lenguaje en los seres humanos: el área de Broca (F5 en los chimpancés) y el área de Wernicke. En el área F5 se han localizado neuronas espejo. Las neuronas espejo se toman a menudo para ser la base de la imitación, pero los primates no humanos son imitadores pobres.
- Comparando los cerebros de humanos y macacos, Liping Wang et al (2015) han localizado un circuito recientemente desarrollado que pudo dotar a los seres humanos con una capacidad única para representar secuencias lingüísticas y no lingüísticas de una manera unificada.
- Según Antoine Balzeau et al (2014), debido a que el sustrato anatómico relacionado con la asimetría del cerebro ha estado presente desde la aparición del linaje de los homínidos (el último ancestro común de chimpancés y el humano moderno tenía ya un cerebro asimétrico), no es posible demostrar una relación directa entre el grado de variaciones en el tamaño, forma, y asimetrías de la tercera circunvolución frontal y el origen de lenguaje.
Michael C. Corballis (2013) repasa las cuestiones probadas referentes a la lateralidad.
- La asimetría es innata.
- La lateralidad y la asimetría se han detectado en el feto y alrededor de dos tercios de los casos muestran el sesgo hacia la izquierda. En la décima semana de gestación, la mayoría de los fetos mueven el brazo derecho más que el izquierdo en la 15 ª, chupan más el pulgar derecho que el izquierdo. A los tres meses, la mayoría de los fetos muestra una ampliación hacia la izquierda del plexo coroideo. Esta asimetría puede estar relacionada con la ampliación hacia la izquierda del plano temporal (parte del área de Wernicke), evidente a las 31 semanas.
- La asimetría se manifiesta de distinto modo en los individuos.
- El 70% de los zurdos utilizan el hemisferio izquierdo para el lenguaje.
- Los genes influencian pero no determinan la lateralidad.
- Un 23% por ciento de los gemelos monocigóticos, que comparten los mismos genes, utilizan preferentemente la mano opuesta (gemelos espejo). En la mayoría de estos gemelos el hemisferio izquierdo es dominante para el lenguaje. En los gemelos, como en los nacidos por separado, se estima que sólo alrededor de una cuarta parte de la variación en el uso de las manos se debe a influencias genéticas.
- Ser zurdo surge de la falta de un sesgo genético hacia la mano derecha, y es cuestión de azar.
- Varios estudios de todo el genoma a gran escala han fracasado en revelar un locus único asociado significativamente con el uso de las manos. Parece que la lateralidad es un rasgo poligénico en parte controlado por los genes que establecen la asimetría del cuerpo en los momentos tempranos del desarrollo.
- Un estudio ha demostrado que las repeticiones de una secuencia de CAG en el locus del receptor del andrógeno en el cromosoma X están relacionadas con la lateralidad. En las hembras, la incidencia de zurdas aumentó con el número de repeticiones, mientras que en los varones se reduce con el número de repeticiones. Este hallazgo apoya un papel de la testosterona en la determinación de la lateralidad.
- En formulaciones recientes de la teoría X-Y, se ha propuesto que la asimetría cerebral está universalmente codificada en el genoma humano, y que las variaciones que dan lugar a la esquizofrenia o anomalías de lateralidad y asimetría cerebral son epigenéticas. Parece que el cambio epigenético a través de la metilación del ADN puede transmitirse entre generaciones.
- Los zurdos no están discapacitados.
- La insuficiencia de la dominancia cerebral (ambidiestros) está asociada con la tartamudez y el déficit de habilidades académicas, dificultades en la salud mental y esquizofrenia. La dominancia invertida (zurdos) no está asociada con déficit alguno. Por el contrario, personas zurdas destacan en todos los ámbitos de la vida.
Si damos por buena una coevolución entre el lenguaje y la producción de herramientas (Corballis, 2003; Hopkins et al., 2007; Sherwood et al., 2007; Hopkins y Nir, 2010; Stout y Chaminade, 2012; Hopkins y Cantalupo, 2004; Stout et al, 2015), cualquier dato acerca de la aparición de la tendencia a utilizar la mano derecha se convierte en una evidencia acerca de la lateralización en su conjunto.
- Mediante el estudio de las marcas dejadas en las esquirlas obtenidas al fabricar las herramientas líticas, Toth (1985) concluyó que la tendencia a utilizar la mano derecha está presente en la cultura olduvaiense hace 2 Ma. Toth y Schick (1993) sugieren una relación con el desarrollo de las áreas lingüísticas.
- El sentido de los modelos que ligan lenguaje, destreza y lateralización cerebral puede someterse a prueba mediante el examen de otros primates. Se ha seguido tradicionalmente la tesis de Warren (1980) de que la lateralización es un fenómeno que solo aparece en nuestra especie.
- Ernest Tabiowo y Gillian S. Forrester (2013) han observado el uso de las manos en acciones espontáneas por parte de 13 gorilas occidentales de tierras bajas cautivos (Gorilla gorilla gorilla). Los resultados muestran un sesgo significativo en el uso de la mano derecha. Según los autores, estos hallazgos sugieren que la dominancia de la diestra es un rasgo de comportamiento relacionado con el predominio del hemisferio izquierdo para el procesamiento de secuencias estructuradas de acciones, y fue heredado por un ancestro común de los seres humanos y los simios.
- Los chimpancés no muestran preferencia por el uso de una mano en particular. Sin embargo, puede constatarse un predominio del planum temporale del hemisferio izquierdo, un componente esencial del área de Wernicke de los humanos (Gannon, 1998). En el chimpancé, el área de Broca, situada en el hemisferio izquierdo, tiene la misión de controlar los movimientos de la mano. Partiendo de la hipótesis de una asimetría fijada por los antecesores comunes de humanos y chimpancés se pueden seguir cuatro diferentes líneas de interpretación respecto a la intervención de la lateralización en el desarrollo del lenguaje.
- Sostener que esta asimetría de los antecesores no tuvo ninguna función comunicativa. Esta función aparecería más tarde solo en los humanos. La función que tiene en los chimpancés es desconocida.
- La asimetría tuvo funciones de comunicación. En los chimpancés permite una comunicación visual basada en gestos.
- El planum temporale izquierdo carece de función comunicativa, antigua o moderna.
- Se trata de una homoplasia. Esta es la menos parsimoniosa de las interpretaciones.
- Merchant y McGrew (1998) señalan que los chimpancés cautivos parecen comportarse como si existiera preferencia por la diestra mientras que los que permanecen en su hábitat original solo muestran esta preferencia en el manejo de herramientas. Indican que una parte de nuestras convicciones acerca de la lateralización y su filogénesis puede basarse en supuestos que no están bastante contrastados.
Peña-Melián, Antonio Rosas et al analizaron tres cráneos fósiles de Homo neanderthalensis encontrados en el yacimiento de El Sidrón (Asturias) (2011) encontrando que el cerebro de los neandertales era asimétrico, más que el del ser humano actual.
Según Natalie T. Uomini (2011), los neandertales producían herramientas de piedra y hueso para ser usadas con la mano derecha, retocaban asimétricamente los artefactos, sus miembros superiores eran muy asimétricos y sus estriaciones dentales son consistentes con el uso de la mano derecha (Almudena Estalrrich y Antonio Rosas, 2013).
Reducción reciente del tamaño del cerebro
Son varias las hipótesis para explicar esta reducción:
- A medida que las sociedades se vuelven más complejas el tamaño del cerebro se reduce, porque aquellos individuos de la especie, que antes no hubieran podido sobrevivir por cuenta propia, ahora logran hacerlo con la ayuda de los otros.
- La reducción puede estar relacionada con la mayor eficiencia de nuestro cerebro.
- Los animales domesticados tienen cabezas más pequeñas que las de sus ancestros salvajes.
El cerebelo
El cerebelo humano, pese a que no ha aumentado tanto de tamaño como el cerebro, contiene el cuádruple de neuronas que el neocórtex y está atrayendo cada vez más atención por su amplia gama de funciones cognitivas.
Robert A. Barton y Chris Venditti (2014) han estudiado la evolución del cerebelo a lo largo de la filogenia de los hominoideos. De acuerdo con los resultados, el cerebelo aumentó rápidamente de tamaño a lo largo de la evolución de los simios, incluyendo los humanos. El aumento de tamaño fue mayor en el cerebelo que en el neocórtex. La aceleración de la expansión del tamaño del cerebelo comenzó en el origen de los simios. El desencadenante de esta expansión pudo ser la forma en que los grandes primates se desplazan bajo las ramas de los árboles de los bosques, con mayor prudencia que los pequeños y planificando sus rutas. Estos resultados sugieren que, en la evolución del cerebro humano, la especialización del cerebelo fue un componente mucho más importante de lo que hasta ahora se ha reconocido y que la inteligencia técnica probablemente haya sido por lo menos tan importante como la inteligencia social en la evolución cognitiva humana. Dado el papel del cerebelo en el control sensorial-motor y en el aprendizaje de secuencias de acción complejas, la especialización cerebelosa es probable que haya apoyado la evolución de avanzadas capacidades tecnológicas de los humanos, que a su vez puede haber sido una preadaptación para el lenguaje.Con respecto al resto de los simios, en nuestra especie el cerebelo es relativamente más pequeño en nuestro género. De acuerdo con Weaver en un estudio que incluyó al H. habilis, H. erectus, neandertales y H. sapiens antiguos y modernos, si bien existe una tendencia a disminuir el tamaño relativo del cerebelo, este patrón cambia súbitamente al llegar al H. sapiens moderno y el cerebelo se hace más grande. Esto explicaría las notables capacidades cognitivas del H. sapiens moderno, en comparación no solo con otros miembros de nuestro género, sino con otros miembros más antiguos de nuestra propia especie.
 |
| Aparato fonador del chimpancé y del HAM. |
La evolución de los órganos relacionados con el lenguaje
Filogénesis del conducto vocal supralaríngeo
- Para Aiello, la posición baja de la laringe es una consecuencia de las adaptaciones anatómicas necesarias para la bipedación. Otra consecuencia de la bipedación es la liberación de la laringe de las tareas locomotrices que desempeña en los simios que la utilizan para dar firmeza a pecho y brazos. Al quedar liberada la laringe de esas funciones, las cuerdas vocales pudieron hacerse más membranosas y se capacitaron así para emitir una mayor variedad de sonidos.
- Laitman (1984) sostiene que los australopitecinos disponían de una laringe en posición alta. El descenso comenzaría en erectus.
- Lieberman, a partir de las marcas dejadas por los músculos, ha afirmado que el habla sería un fenómeno muy tardío, propio de los seres humanos de aspecto moderno y, en parte, de los neandertales. Estos últimos serían capaces de emitir parte de los sonidos al alcance de los humanos, pero no todos (como a, i y u).
- En los neandertales no se observa el descenso de la laringe. Por el contrario, el hioides es idéntico al del sapiens. El hallazgo en Kebara (Oriente Próximo; Arensburg et al, 1989) de un hueso hioides fosilizado (un hueso intermedio entre la laringe y la base del cráneo que da lugar a gran parte de la musculatura lingual y laríngea), perteneciente a un Homo neanderthalensis, nos permite sostener que su aparato fonador era similar al nuestro. Lieberman ha criticado la identificación entre hueso hioides y capacidad de habla.
- Según Duchin (1990) las dimensiones de la cavidad oral de los neandertales no difieren significativamente de la de los humanos modernos.
- Krantz (1988) piensa que un descenso laríngeo completo sólo se realizó hace 40.000 años (¡coincide con la explosión simbólico-artística del Paleolítico!). La primera fase, hace 200.000 años, habría modernizado el cráneo y especialmente la laringe, dando lugar a una cavidad que supondría la mitad de la actual y permitiría una conducta vocal imperfecta.
 |
Audiograma para chimpancés, humanos modernos y primeros homínidos de 0,5-5,0 kHz. Los resultados para P. robustus y A. africanus, son similares, lo que sugiere un patrón y unas capacidades auditivas similares.
(A) La banda ocupada es similar en los chimpancés y los primeros homínidos, pero se desplaza hacia frecuencias ligeramente superiores en los segundos. Los seres humanos modernos muestran una banda que se extiende más hacia frecuencias más altas.
(B) Curvas de transmisión de potencia en decibelios a la entrada de la cóclea para una intensidad plana de onda incidente. plana.La transmisión de potencia de sonido curvas corresponden a decibelios en la entrada a la cóclea en relación con P 0 = 10 -18 W para una intensidad de la onda plana incidente de 10 elevado a -12 W/m2. Los puntos más altos a lo largo de la curva indican mayor volumen transmitido y mayor sensibilidad auditiva.
|
Filogénesis del oído
La comunicación oral humana precisa tanto la capacidad de articular un amplia gama de sonidos como la de oírlos, es decir, una alta sensibilidad adaptada a un rango amplio de frecuencias. En los audiogramas, los HAM se caracterizan por una amplia región de mayor sensibilidad entre aproximadamente 1,0 y 4,5 kHz, mientras que los chimpancés muestran una disminución de la sensibilidad por encima de 3,0 kHz y alcanzan un mínimo entre 4,0 y 5,0 kHz. Entre 1,0 y 5,0 kHz, los valores medios de transmisión de potencia de sonido son significativamente más altos en el HAM que en los chimpancés, excepto entre 2,0-2,5 kHz. Del mismo modo, la banda ocupada en los seres humanos es significativamente más amplia (ca. 43%) que en los chimpancés y se desplaza hacia frecuencias más altas. Consonantes como la t, k, f, y s tienen una cantidad considerable de energía concentrada dentro de la gama de frecuencias de 3 a 5 kHz. De hecho, el uso de consonantes es una de las principales distinciones, junto con un componente simbólico, entre el lenguaje humano y la mayoría de las formas de comunicación animal.
Las llamadas de baja frecuencia, generalmente ≤1 kHz, emitidas por los primates arborícolas, viajan lejos en el dosel del bosque lluvioso. Sin embargo, la atenuación de la señal a distancias ≥25 m es considerablemente mayor en entornos abiertos, tales como la sabana. Por el contrario, la comunicación de corto alcance es más fuertemente dependiente del ruido ambiental de fondo, que es más bajo en la sabana en la mayoría de frecuencias de hasta 10 kHz. Esta combinación de una mayor atenuación de la señal y menor ruido ambiental de fondo implica que en hábitats abiertos se ve favorecida la comunicación intragrupo de corto alcance y se puede esperar que la señal sea más elaborada y variada.
Las llamadas de baja frecuencia, generalmente ≤1 kHz, emitidas por los primates arborícolas, viajan lejos en el dosel del bosque lluvioso. Sin embargo, la atenuación de la señal a distancias ≥25 m es considerablemente mayor en entornos abiertos, tales como la sabana. Por el contrario, la comunicación de corto alcance es más fuertemente dependiente del ruido ambiental de fondo, que es más bajo en la sabana en la mayoría de frecuencias de hasta 10 kHz. Esta combinación de una mayor atenuación de la señal y menor ruido ambiental de fondo implica que en hábitats abiertos se ve favorecida la comunicación intragrupo de corto alcance y se puede esperar que la señal sea más elaborada y variada.
Ignacio Martínez et al analizaron 5 cráneos procedentes de la Sima de los Huesos y llegaron a la conclusión de que manifiestan una capacidad auditiva muy similar a la de la humanidad actual, con una banda de alta sensibilidad entre 1-5 kilohercios, aunque con un límite superior ligeramente más bajo.
Según Rolf M. Quam et al (2013) el martillo y el tímpano evolucionaron temprano y las características actuales ya estaban presentes en Australopithecus africanus. Los primeros homínidos parecen estar caracterizados por un martillo de tipo humano, mientras que el yunque y el estribo son primitivos y más similares a los de los chimpancés en su tamaño y forma.
Según Rolf M. Quam et al (2013) el martillo y el tímpano evolucionaron temprano y las características actuales ya estaban presentes en Australopithecus africanus. Los primeros homínidos parecen estar caracterizados por un martillo de tipo humano, mientras que el yunque y el estribo son primitivos y más similares a los de los chimpancés en su tamaño y forma.
Rolf Quam et al (2015) han estudiado la anatomía del oído externo y medio y estimado las capacidades auditivas de Australopithecus africanus y Paranthropus robustus.
- En comparación con los chimpancés, estos taxones son más derivados en su canal auditivo externo un poco más corto y más ancho, membrana timpánica más pequeña, y relación de palanca de martillo/yunque inferior.
- Siguen siendo primitivos en el pequeño tamaño de la base del estribo.
- El volumen de la cavidad timpánica y los tamaños de la membrana timpánica y base del estribo, parecen ser ligeramente menores en P. robustus que en A. africanus.
- En comparación con los chimpancés, los dos taxones homínidos tempranos muestran mayor sensibilidad que chimpancés y HAM a las frecuencias entre 1,5 y 3,5 kHz. Por encima de 3,5 kHz, los primeros homínidos son menos sensibles que los humanos modernos, pero similares a los chimpancés. La banda ocupada es similar a la de los chimpancés, pero desplazada hacia frecuencias más altas en ambos extremos superior e inferior.
Evolución de características filogenéticas relacionadas con el comportamiento social
El enfoque neuroetológico, predice especializaciones biológicas para la vida social en primates humanos y no humanos.
La evidencia revisada indica que los mecanismos ancestrales a menudo se han duplicado, reutilizado, o regulado para apoyo al comportamiento social. Centrándose en la investigación reciente de primates no humanos, el cerebro pudo implementar funciones sociales por cooptación y ampliación de los mecanismos ya existentes que anteriormente apoyaban funciones no sociales.
Las diferencias en el comportamiento social entre las especies, así como entre los individuos dentro de una especie, son consecuencia en parte de la regulación neuromoduladora de circuitos neuronales, que a su vez parecen estar bajo control genético parcial. En última instancia, la variación intraespecífica en el comportamiento social tiene consecuencias diferenciales de aptitud, proporcionando los bloques fundamentales de construcción de la selección natural (Steve W.C. Changa et al, 2013)Amy L. Bauernfeind et al (2013) estudiaron la corteza insular en 30 especies de primates, encontrando que en sapiens se ha producido un crecimiento extraalométrico, proporcionando la base para la mejora de la cognición social y que, en relación al chimpancé, es una de las áreas cerebrales más ampliadas.
Los humanos somos los únicos primates con la esclerótica ocular blanca. El cerebro detecta automática y rápidamente cualquier señal procedente del contraste del irirs y el blanco de los ojos de otros, informándonos de sus intereses y emociones.
Otra importante característica relacionada con la socialidad es la riqueza de las expresiones faciales, derivada de la presencia de 42 músculos.
Para C. Crockford et al (2013), la evolución de la inteligencia social compleja está relacionada con cambios en el sistema hormonal que favorecieron la producción de oxitocina.
Evolución de características filogenéticas relacionadas con la dieta
Las proporciones entre la longitud del intestino y la del cuerpo y entre la superficie gastrointestinal y corporal, son más similares a las de los carnívoros que a las del resto de los primates.
Al contrario que en el resto de los primates, y tal como sucede en los carnívoros, el ser humano asimila mejor el hierro de la hemoglobina que el procedente de los vegetales.
Los humanos somos hospedadores definitivos de diversas especies de cestodos del género Taenia, que parasitan también a los grandes carnívoros. Del análisis genético de estas especies se deduce que su adaptación a infectar a los humanos debió suceder muy pronto, en cuanto pasamos a una dieta más carnívora, y antes de la utilización del fuego, quizá hace 2,5 Ma, coincidiendo con la aparición del género Homo.
Karen R. Rosenberg and Wenda R. Trevathan, Nov. 2001. The Evolution of Human Birth, Scientific American.
Evolución y genómica del cerebro humano. 2018. M.A.Rosales-Reynoso, C.I.Juárez-Vázquez, P. Barros-Núñez
Posible relación entre el cerebro y la piel y el pelo
A medida que los humanos ancestrales cambiaban su hábitat de los bosques tropicales hacia las sabanas abiertas, la pérdida de su vello corporal facilitaba la disipación del calor corporal. La exposición de la piel a los estímulos externos pudo estimular un aumento en el tamaño del cerebro para procesar centralmente una gran cantidad de nueva información sensorial. Varias líneas de evidencia apoyan esta hipótesis:- El origen embriológico común de la epidermis y el sistema nervioso central (SNC) del neuroectodermo primitivo, dotó a la piel de un amplio conjunto de mecanismos neurológicos que reflejan el SNC. La desnudez humana también estimuló nuevas respuestas psicosociales y de estrés, mediadas por la oxitocina cutánea y los glucocorticoides, respectivamente.
- Dos grupos de organismos marinos, los cefalópodos y los peces eléctricos, muestran una gran proporción de masa cerebral asociada con capacidades sensoriales cutáneas aumentadas.
Enlaces:
La evolución del sentido del oído humano. Entrevista a Ignacio Martínez Mendizábal.Karen R. Rosenberg and Wenda R. Trevathan, Nov. 2001. The Evolution of Human Birth, Scientific American.
Evolución y genómica del cerebro humano. 2018. M.A.Rosales-Reynoso, C.I.Juárez-Vázquez, P. Barros-Núñez
Te puede interesar:
|
No hay comentarios:
Publicar un comentario