 |
| (a) Patrón filogenético a través del tiempo, con repetidas expansiones, extinciones y asimilaciones. (b) Expresión del patrón anterior en el registro fósil y arqueológicos. (c) Fósiles clave y yacimientos arqueológicos africanos entre hace 400-130 ka, en el contexto del cambio climático. Las líneas azules representan fósiles de Homo heidelbergensis, los círculos y líneas moradas Homo helmei y los círculos y líneas rojas Homo sapiens. Los yacimientos arqueológicos están representados por líneas negras. Las industrias achelenses, por un bifaz y las industrias de la MSA por una punta Levallois. Una nueva datación ha retrasado Jebel Irhoud hasta hace 315 ka. Mirazón, 2016. |
 |
| Mandíbula de Misliya. Israel Hershkovitz et al, 2018 |
En 1969 Blumenbach proporcionó el primer diagnóstico morfológico de Homo sapiens: barbilla, mandíbula pequeña, caninos e incisivos pequeños, caja craneal grande sobre una columna vertebral vertical, postura erguida, bipedación, pelvis cóncava, nalgas y pulgar largo.
Los especímenes atribuidos a sapiens muestran entre sí profundas diferencias morfológicas que han dado lugar a la distinción entre arcaico y anatómicamente moderno (Trinkaus, 1982). Estos últimos han sido caracterizados de forma imprecisa por un esqueleto más grácil, un cráneo más redondeado y cara, mandíbula y dientes más pequeños. La atribución de los fósiles a arcaicos o modernos es caprichosa y sin base morfológica.
De acuerdo con Lieberman (1998) el cráneo de un HAM debe mostrar:
- Una caja craneal globular.
- Frente vertical.
- Arcos supraciliares poco marcados.
- Fosa canina.
- Mentón pronunciado (James D. Pampush y David J. Daegling, 2016).
 |
| A la izquierda, reconstrucción basada en los ejemplares de Jebel Irhoud. A la derecha, humano moderno. NHM Londres. |
Especímenes más antiguos que exhiben algunas de estas características:
Christopher Brian Stringer y Laura Tabitha Buck (2014) consideran que los especímenes de Homo sapiens muestran rasgos distintivos en la morfología del cráneo, el oído interno, la dentición y la pelvis. Además, la genética, puede proporcionar datos complementarios y tal vez incluso más fructíferos para caracterizar las diferencias entre el sapiens y sus parientes cercanos.- La edad de 0,64 Ma atribuida a los especímenes de Bodo llevaría el origen de nuestra especie lejos del rango en que se mueven los indicios proporcionados por los métodos moleculares, si es que el “hombre de Bodo” debe considerarse ya como un ser humano de aspecto moderno. Y de otorgar esa condición al ejemplar de Danakil, entonces nos habríamos remontado a cifras que rondan el millón de años.
- Jebel Irhoud, 55 km al sureste de Safi, Marruecos (Hublin et al, 2017). En 1961 se halló accidentalmente un cráneo casi completo, Irhoud 1, lo que promovió excavaciones dirigidas por Émile Ennouchi (1962) y Jacques Tixier y Roger de Bayle des Hermens (1967 y 1969) que proporcionaron diversos fósiles humanos asociados con restos de fauna y herramientas levallois. Solo se registró con precisión la localización de un húmero y la datación resultó dudosa. Nuevas excavaciones más recientes (Hublin et al, 2017) permitieron la recuperación in situ de restos humanos. Es probable que todos o la mayoría de los restos humanos del yacimiento fuesen acumulados en una estrecha ventana temporal, correspondiendo con la formación de la capa 7. Los restos corresponderían a tres adultos, un adolescente y un individuo de unos 7,5 años de edad. El cerebro es más alargado, menos globular que en el humano moderno, pero la cara y los dientes son más similares. Fueron datados en hace 315± 34 ka por termoluminiscencia y U-series (Richter et al, 2017). Para el paleoentorno, ver Geraads et al, 2013; hace 300 ka el Sahara estaba repleto de ríos.
- Irhoud 1. Mandíbula. 1961. Depósitos inferiores.
- Irhoud 2. Calota. 1962. Depósitos inferiores.
- Irhoud 3. Mandíbula de juvenil. 1968. Depósitos inferiores.
- Irhoud 4. Húmero juvenil. 1969. Capa 18 de Tixier.
- Irhoud 5. Coxal juvenil. 1969. Capa 18 de Tixier.
- Irhoud 6. Fragmento de mandíbula. 1961-1969. Identificado entre restos de fauna.
- Irhoud 7. Primer premolar inferior derecho. 2004. Limpieza inicial.
- Irhoud 8. Parte distal de un molar inferior izquierdo. 2004. Limpieza inicial.
- Irhoud 9. Primer o segundo molar inferior. 2006. Capa 4.
- Irhoud 10. Cráneo. 2007. Capa 7.
- Irhoud 11. Mandíbula. 2007. Capa 7.
- Irhoud 12. Incisivo inferior. 2007. Capa 7.
- Irhoud 13. Fragmento proximal de un fémur izquierdo. 2007. Capa 7.
- Irhoud 14. Costilla. 2009. Capa 7.
- Irhoud 15. Costilla. 2009. Capa 7.
- Irhoud 16. Húmero juvenil. 2009. Capa 7.
- Irhoud 17. Fragmento proximal de un fémur derecho. 2009. Capa 7.
- Irhoud 18. Vértebra lumbar. 2007. Limpieza inicial.
- Irhoud 19. Peroné. 2009. Capa 7.
- Irhoud 20. Vértebra cervical. 2009. Limpieza inicial.
- Irhoud 21. Maxilar. 2011. Capa A.
- Irhoud 22. Segundo y tercer molares superiores derechos. 2011. Capa A.
- Homo helmei y Homo sapiens idaltu
- En Kenia (Koobi Fora) se han encontrado restos humanos cercanos a los humanos de aspecto moderno, con una antigüedad de 300-270 ka obtenida mediante series de uranio.
- Etiopía (Collard y Dembo, 2013). Los más importantes son Omo I, Omo II y Omo III, datados en hace 195 ka y Herto, datado hace 160 ka.
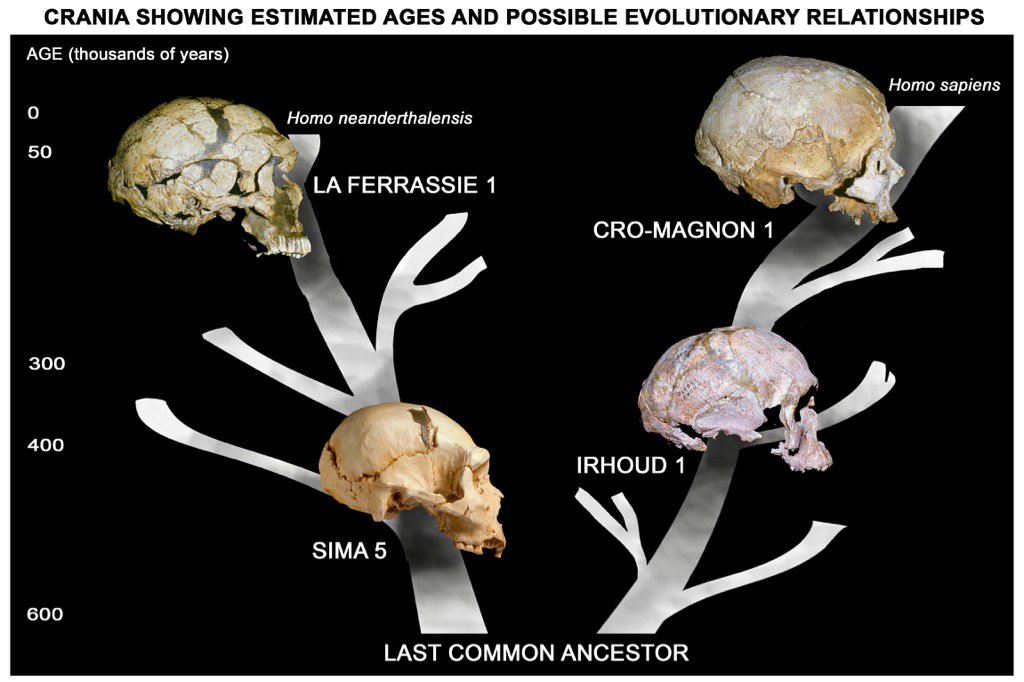 |
| Posibles relaciones evolutivas de algunos ejemplares craneales. Chris Stringer. 2017. |
- Stringer (1974, 1978) encontró que ciertos ángulos e índices craneales pueden ser usados para separar a los HAM, los sapiens del PS y fósiles como Skhul y Omo Kibish 1 de neandertales y otros grupos arcaicos. Homo sapiens se caracterizaría por un neurocráneo en cúpula, reducción en el tamaño y proyección de la cara y un aumento de la flexión craneoespinal.
- Day y Stringer (1982) elaboraron una lista de características de diagnóstico y las utilizaron para incluir a Omo 1 en Homo sapiens sensu stricto, pero no a Omo 2. Según ellos, para asignar un fósil a sapiens debería mostrar al menos un 50% de las características. Estudios posteriores con poblaciones actuales y fósiles hacen descartar la utilidad de la lista.
- Problemas de definición:
- La variación entre Omo 1 y Omo 2 puede reflejar la coexistencia de dos poblaciones o que la relación entre la morfología de heidelbergensis y la de sapiens no es simple y lineal.
- Según el Modelo Multirregional Africano, existieron diferencias regionales dentro del continente, con diferentes poblaciones interfértiles (Stringer, 2003, 2012). Las fluctuaciones climáticas entre glaciales/interglaciales y ciclos climáticos cortos pudieron crear/remover las barreras biogeográficas originando la contracción de las poblaciones y la expansión y contacto en los momentos favorables (Blome et al, 2012; Coulthard et al, 2013; Ziegler, 2013; Hublin et al, 2017).
- El hallazgo de especímenes relativamente recientes con rasgos arcaicos (Iwo Eleru, Ishango) ha sido interpretado como un resultado de la hibridación entre HAM y especies arcaicas, quizás Homo heidelbergensis. Hammer et al (2011) sugieren una introgresión en África Central de una rama que se separó de la del HAM hace ca 700 ka. Mendez et al (2013) estimaron una fecha de coalescencia de 581 ka para el cromosoma Y, fuera del rango de la datación de los fósiles de sapiens más antiguos conocidos, lo que puede reflejar una hibridacion con especies extintas.
- La pelvis de Omo 1 sugiere que se trata de una hembra (Ashley S. Hammond, Danielle F. Royer y John G. Fleagle, 2017)
- Por otra parte, se ha constatado la hibridación del sapiens en Eurasia con neandertales y denisovanos.
- Para los autores, el grado de introgresión es pequeño y no afecta a las carcterísticas morfológicas del sapiens.
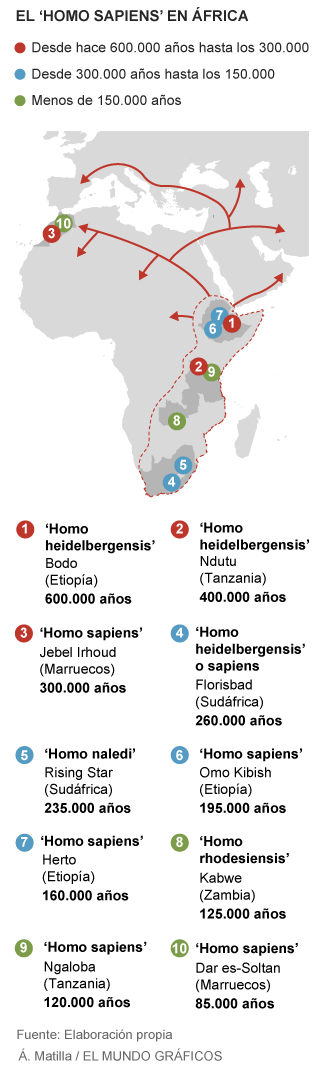 |
| El Homo sapiens en África. El Mundo. |
Jeffrey H. Schwartz (2016) resume los problemas de clasificación.
- Barbilla. Además de los especímenes del PS, como Abri Pataud, Grimaldi, Dolni Vestonice, Predmosti, Ohalo, e Isturitz, solo Tianyuan PA 1281, KRM 41815, y Tuinplaas presentan barbilla.
- Las regiones sinfisarias de Fish Hoek, Boskop y Klasies River Mouth KRM 13400 y 14695 son lisas con abultamiento variable. Solo KRM 41815 parece presentar una quilla central; inferiormente su región sinfisaria no es significativamente más gruesa que el hueso lateral.
- La línea media de la mandíbula de Zhirendong, carece de quilla y presenta una mínima protuberancia más pronunciada muy por encima del margen inferior.
- La región sinfisaria de la mandíbula de Tuinplaas presenta una T invertida, pero, inferiormente, no es apreciablemente más gruesa que el hueso lateral (Schwartz y Tattersall, 2000).
- Prominencia supraorbital dividida (Stringer et al, 1984).
- Los fósiles clasificados como arcaicos presentan una protuberancia supraorbital continua a través de la glabela, con márgenes gruesos y redondeados (Eyasi, Eliye Springs, Omo Kibish, Guomde, Laetoli (LH18), y Ndutu (Este de África), Florisbad (Sur de África), Jebel Irhoud (pero no Irhoud 2, que muestra una separación clara) y Dar es-Soltane (Norte de África), Dali y Jinniushan (China) y Narmada (India) (Schwartz y Tattersall, 2003).
- Entre los fósiles clasificados como modernos, las prominencias supraorbitales de Border Cave, Qafzeh 6, y todos los ejemplares adultos de Skhul, son más bien lisas a través de la glabela, con márgenes redondeados (Schwartz y Tattersall, 2003). Herto (BOU-VP-16/1), asignado a Homo sapiens idaltu, fue descrito con prominencia supraorbital dividida (White et al, 2003), sin embargo medialmente y en cada extremo, la prominencia supraorbital es alta, plana, orientada lateral oblicuamente y circunscrita supero medialmente por un borde bien definido.
- Los especímenes europeos del PS presentan una prominencia supraorbital que ha sido descrita como dividida en algún caso con benevolencia (e.g. Abri Pataud) o justamente (e.g. Mladeč). También presenta una prominencia dividida el cráneo parcial de Tuinplaas.
- Los especímenes con una verdadera barbilla deberían ser considerados Homo sapiens.
- Combinaciones de una cara relativamente pequeña y sin prognatismo y un cráneo alto y abovedado, sin una verdadera barbilla (por ejemplo LL1, LL2, Qafzeh 6 y 7, Fish Hoek, Boskop, BOU-VP-16/1), representan morfos estrechamente relacionados con Homo sapiens.
- Un cráneo alto y abovedado y una cara prognática sin una verdadera barbilla (por ejemplo Skhul V), pertrenece a un morfo hermano del grupo anterior.
Te puede interesar:
|
No hay comentarios:
Publicar un comentario