 |
| Cráneo MRD-VP-1/1. Foto de Dale Omori. Cleveland Museum of Natural History |
Australopithecus anamensis M. Leakey et al (1995).
- Holotipo. KNM-KP 29281. Mandíbula inferior con todos los dientes, en la que falta la rama ascendente. 4 Ma. Descubierta por Peter Nzube (1994). Fragmento de hueso temporal asociado. Perteneció a un individuo femenino de pequeño tamaño. Se asemeja, por su tamaño y su ancho, a la de los monos del Mioceno. Los dientes están más alineados y paralelos que en A. afarensis. En vista lateral el frente de la mandíbula se encuentra marcadamente retraído y en vista oclusal (desde arriba) se observa un torus interno muy desarrollado y elongado posteriormente.
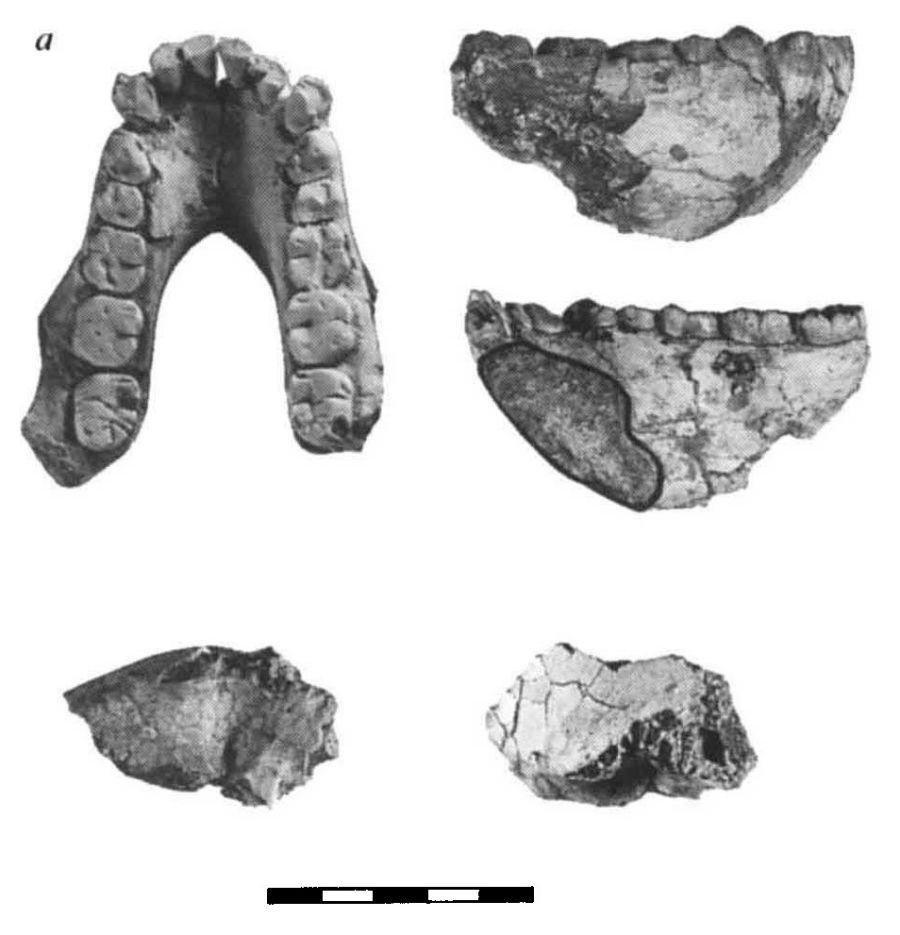 |
| KNM-KP 29281, mandíbula en vista oclusal (superior izquierda), lateral derecha (superior derecha) y medial (centro a la derecha). En la parte inferior, el temporal en vista inferior (abajo a la izquierda) y lateral (abajo a la derecha). (Leakey, et al. 1995) |
- Yacimientos.
- Kanapoi, entre los ríos Kalabata y Kakurio, suroeste del Lago Turkana, Koobi Fora, Norte de Kenia.
- Allia Bay, este del Lago Turkana.
- Woranso-Mille (WORMIL), Afar. Etiopía. Distintas áreas: Am-Ado (AMA), Aralee Issie (ARI), Mesgid Dora (MSD) Makah Mera (MKM) y Miro Dora (MRD).
- Middle Awash, nordeste de Etiopía.
- Se ha reportado la presencia en Ngorongoro (Tanzania) de huellas de un ser bípedo, datadas en hace 3,7 Ma.
4,19-3,72 Ma.
Anam significa lago
en la lengua de Turkana.
El primer fósil
correspondiente a esta especie fue encontrado en 1965 por Bryan Patersson en la
localidad de Kanapoi (Kenia).
Los hallazgos sugieren la existencia de al menos cuatro poblaciones sucesivas en el tiempo pero alopátridas (Woranso-Mille, Allia Bay, Asa Issie y Kanapoi) con morfologías craneal y dental diferenciadas (Haile-Selassie et al, 2019).
Los hallazgos sugieren la existencia de al menos cuatro poblaciones sucesivas en el tiempo pero alopátridas (Woranso-Mille, Allia Bay, Asa Issie y Kanapoi) con morfologías craneal y dental diferenciadas (Haile-Selassie et al, 2019).
Hallazgos
Kanapoi
56 fósiles de 47 individuos (4 mandíbulas de adulto, un maxilar de adulto y dientes deciduos y permanentes; fragmento de húmero, capitate, fragmento proximal de falange de la mano y porciones proximal y distal asociadas de una tibia).- KNM-KP 271 Homínido de Kanapoi. Primer resto descubierto de la especie, por Bryan Patterson en 1965. Fragmento distal de húmero izquierdo de 4 Ma. Se asemeja morfológicamente al de Homo por lo que se dudó en su asignación.
- KNM-KP 29281 Holotipo.
- KNM-KP 29282 LM1 o M2.
- KNM-KP 29283 Maxilar. Las grandes raíces de los caninos condicionan la morfología alrededor de los orificios nasales.
- KNM-KP 29284 RC & RP3 germinales
- KNM-KP 29285 Descubierto por Kamoya Kimeu. (1994). Tibia a la que le ha desaparecido su porción media de 4,1 Ma. Claros indicios de bipedación, la más temprana evidencia en los australopitecinos. Los cóndilos proximales de la tibia son cóncavos como en los humanos. La porción distal de la tibia presenta un engrosamiento óseo que actuaría como una estructura que absorbería el impacto de la locomoción bípeda.
- KNM-KP 29286 Fragmentos de mandíbula y dentición asociada (RI1, L & RI2-M3)
- KNM-KP 29287 Mandíbula sin dientes.
- KNM-KP 30498 L & R fragmentos maxilares y dentición asociada.
- KNM-KP 30500 Fragmentos mandibulares y dentición asociada.
- KNM-KP 30502 Fragmentos de dientes asociados a mandíbula.
- KNM-KP 30503 Falange proximal de una mano.
- KNM-KP 30505 Parcial M germinal.
- KNM-KP 30942 5 fragmentos de diente.
- KNM-KP 31712 Mandíbula juvenil y fragmentos dentales asociados. 2,8 años en el momento de la muerte (Smith et al, 2015).
- KNM-KP 31713 Mandíbula con fragmentos de diente.
- KNM-KP 31714 Ldm2.
- KNM-KP 31715 LM1 o M2 fragmentario y otros dos fragmentos de diente.
- KNM-KP 31716 P3 o P4 fragmento y C/ fragmentos.
- KNM-KP 31717 LM3, RM3 & LM2 fragmentos.
- KNM-KP 31718 R Mandibular fragmento (M2–3).
- KNM-KP 31719 I1.
- KNM-KP 31720 Fragmento M maxilar.
- KNM-KP 31721 RM2 & M3 coronas parciales.
- KNM-KP 31723 RM3.
- KNM-KP 31724 L capitate.
- KNM-KP 31726 RP4.
- KNM-KP 31727 RC.
- KNM-KP 31728 LM1.
- KNM-KP 31729 Rdm2.
- KNM-KP 31730 LM2 & RP3.
- KNM-KP 31732 Fragmentos de diente.
- KNM-KP 34725 Fragmentos de cráneo y dentición juvenil asociada. 3,63-4,25 años en el momento de la muerte. (Smith et al, 2015).
- KNM-KP 35838 LM3.
- KNM KP 35839 Asociados LI1, RC & LP3.
- KNM-KP 35840 LM3 & maxilar M fragmentos.
- KNM-KP 35841 M corona.
- KNM-KP 35842 R maxilar M.
- KNM-KP 35844 M fragmento.
- KNM-KP 35845 M fragmento.
- KNM-KP 35847 LM2.
- KNM-KP 35850 Maxilar M fragmento.
- KNM-KP 35851 LM2 o M3 fragmento.
- KNM-KP 35852 LC.
- KNM-KP 37522 L mandibular molar.
- KNM-KP 37523 M fragmento.
- KNM-KP 37524 Fragmentos de diente.
- KNM-KP 47951 – inferior; LC1-P3, RP3–4, dos fragmentos de diente.
- KNM-KP 47952 – superior; LI1−2, LC1, frgmento de RI1
- KNM-KP 47953 – inferior; RC1-M1, RM3, LC1, LM3
- KNM-KP 47954 – superior; RP3
- KNM-KP 47955 – inferior; LM1
- KNM-KP 47956 – Mandíbula parcial izquierda
- KNM-KP 47957 – inferior; LM3 germinal
- KNM-KP 49388 – superior; LP4
- KNM-KP 52120 – superior; LM2
{kind=link}
{kind=link}
Allia Bay
Meave Leakey. 31 fósiles (fragmento mandibular de adulto, dos maxilares fragmentarios de adulto y dientes deciduos y permanentes). Datados en 3,9 Ma.- KNM-ER 7727 LM2
- KNM-ER 20419 L radio
- KNM-ER 20420 LM2
- KNM-ER 20421 RM3
- KNM-ER 20422 LM1
- KNM-ER 20423 LM2
- KNM-ER 20427 LM1
- KNM-ER 20428 LM3
- KNM-ER 20432 L mandibular fragmento (P3–4). P3 y caninos inferiores más parecidos a los de afarensis.
- KNM-ER 22683 LP4
- KNM-ER 24148 Ldm2
- KNM-ER 30200 L maxilar fragmento (M1–2, parxial M3)
- KNM-ER 30201 LM1
- KNM-ER 30202 RIl
- KNM-ER 30731 RC
- KNM-ER 30744 RC
- KNM-ER 30745 L maxilar fragmento (parcial C, P3–M1, parcial M2, M3). Ver vídeo.
- KNM-ER 30747 LP4
- KNM-ER 30748 L maxilar M fragmento
- KNM-ER 30749 19 LM1
- KNM-ER 30750 RC
- KNM-ER 35228 RP4
- KNM-ER 35229 L mandibular M fragmento
- KNM-ER 35230 M fragmento
- KNM-ER 35231 RM1 o M2
- KNM-ER 35232 LM1
- KNM-ER 35233 LM2
- KNM-ER 25234 LP3
- KNM-ER 35235 LM2
- KNM-ER 35236 LM3
- KNM-ER 35238 RM1
Sibilot Hill.
A unos 20 km de Allia Bay. Fragmentos de un radio de adulto, atribuidos a la especie en base a su datación.
 |
| Localización de Woranso-Mille |
Woranso-Mille (WORMIL)
Descubierto en 2004. Estudiado por Haile Selassie. 55 restos, la mayoría dientes aislados. Datado en 3,87-3,7 Ma. La fauna asociada sugiere un ambiente cerrado de bosques de ribera rivera y agua abundante.
Los restos de las extremidades superiores son similares a los de Au. afarensis (con una morfología similar a la de los primates arbóreos), pero la tibia tiene rasgos asociados con la bipedia.
La muestra de Allia Bay es más parecida a afarensis en varios detalles morfológicos, destacando ER 20432.
Los dientes de la muestra de Woranso-Mille muestran un mosaico de caracteres típicos de anamensis junto con otros de afarensis. La dentición es menos primate que la de ramidus. Los P4 mandibulartes son más similares a afaremsis, sin embargo, algunos ejemplares muestran rasgos distintivos en la corona: una esquina distolingual extremadamente abultada, asociada con un talónido relativamente grande y una corona mesiodistalmente alargada. Esta morfología única se acompaña de una forma de la raíz que es diferente de las identificadas previamente (Yohannes Haile-Selassie y Stephanie M. Melillo, 2014).
Son rasgos avanzados el mayor tamaño de los poscaninos, con premolares oclusalmente más complejos y esmalte dental más grueso. Los caninos son más pequeños que los de Ardipithecus ramidus y su desgaste es menos simiesco que en los antiguos homínidos conocidos.
Por la fauna asociada a los hallazgos, sabemos que vivió en un mosaico de hábitats: sabanas arboladas y secas, bosques en galería que bordean los ríos y praderas más abiertas (Ungar et al, 2017). Se trata de un hábitat muy similar al del Australopithecus afarensis.
- MSD-VP-5/16. Mandíbula izquierda bien preservada con M1-2, rota anteriormente al nivel de I2. Fue hallada en 2006. Carece de ramus ascendente. Probablemente perteneció a una hembra de tamaño comparable a A.L. 128-23. M1-2 son grandes en relación al tamaño corporal. Comparte con afarensis una serie de características como un corpus robusto, un hueco en el corpus lateral y una sínfisis más vertical.
- MSD-VP-5/50. Mandíbula izquierda con P3-M3. Preservada parte del ramus ascendente. Hallada en 2009. Menos parecido a afarensis. La altura y robustez del corpus a la altura de M1 están fuera de los rangos tanto de anamensis como de afarensis (algo menos robusta).
- ARI-VP-1/190. Canino inferior deciduo, similar al de afarensis en la morfología de su corona. Raíz más larga y relieve lingual más corto que lo usual en anamensis.
- ARI-VP-2/95. P3 con morfología oclusal similar a afarensis.
- En la localidad de Miro Dora se halló en 2016 un cráneo fósil identificado como MRD-VP-1/1 (Haile Selassie et al, 2019). El primer fragmento –la mandíbula superior– fue hallado por Ali Bereino, un trabajador. Una investigación adicional del área propició la recuperación del resto del cráneo. Se trata del único cráneo conocido de la especie y se ha datado en 3,8 Ma. Los caracteres craneales y el tamaño del canino indican que perteneció a un macho adulto. El yacimiento formaba parte de un pequeño delta de un río parecido al actual río Mille, que también procedía del altiplano etíope, pero que desembocaba en un lago de unos cinco a ocho metros de profundidad, con una cierta salinidad. Los restos de polen y biomarcadores indican que la vegetación era variada. Más lejos debieron de existir hábitats abiertos (Saylor et al, 2019).
Middle Awash
Con una datación de hace ca 4,2 Ma, se recuperaron en Assa Issie, 2ª localidad (2000-2002) los restos axiales de Australopithecus más antiguos conocidos (siete vértebras). El atlas, o primera vértebra cervical (C1), es similar en tamaño a la de Homo sapiens, con sinapomorfias en las facetas articulares y procesos transversales. La ausencia de un tubérculo retroglenoide sugiere que, como en Homo, A. anamensis carecía del músculo atlantoclavicularis, lo que resulta en una capacidad reducida para trepar en comparación con los grandes simios. El T1 es derivado en forma y tamaño, como en Homo, con superficies epifisarias de cuerpo vertebral agrandadas para mitigar las cargas compresivas de gran magnitud de la bipedalidad a tiempo completo. La faceta costal completa es diferente al patrón los grandes simios existentes y representa la evidencia más antigua del patrón univertebral derivado en Hominini. Meyer y Williams, 2019.Otros
La asignación a anamensis de los siguientes restos se debe solo a que su datación es anterior a 3,7 Ma.- BEL-VP-1/1. Frontal de Belohdeli (Asfaw 1987). Datado en hace 3,9 Ma.
- Dientes de Fejej (Fleagle et al 1991; Kappleman et al.1996; Grine et al 2006; Carol V. Ward, 2014). Datados en 4,2-4 Ma.
- Diente y fragmento de fémur de Galili (Haile-Selassie & Asfaw 2000; Macchiarelli et al 2004; Viola et al 2008).
Morfología
Se caracterizan por una presencia simultánea de rasgos primitivos (cráneo) y derivados (poscráneo) que han confirmado que la evolución dentro de los homínidos se produjo durante el Plioceno en forma de mosaico y en todo caso, antes en el aparato locomotor que en el cerebro. La locomoción bípeda se encuentra en un estado incipiente.
Los restos de las extremidades superiores son similares a los de Au. afarensis (con una morfología similar a la de los primates arbóreos), pero la tibia tiene rasgos asociados con la bipedia.
La muestra de Allia Bay es más parecida a afarensis en varios detalles morfológicos, destacando ER 20432.
Los dientes de la muestra de Woranso-Mille muestran un mosaico de caracteres típicos de anamensis junto con otros de afarensis. La dentición es menos primate que la de ramidus. Los P4 mandibulartes son más similares a afaremsis, sin embargo, algunos ejemplares muestran rasgos distintivos en la corona: una esquina distolingual extremadamente abultada, asociada con un talónido relativamente grande y una corona mesiodistalmente alargada. Esta morfología única se acompaña de una forma de la raíz que es diferente de las identificadas previamente (Yohannes Haile-Selassie y Stephanie M. Melillo, 2014).
Son rasgos avanzados el mayor tamaño de los poscaninos, con premolares oclusalmente más complejos y esmalte dental más grueso. Los caninos son más pequeños que los de Ardipithecus ramidus y su desgaste es menos simiesco que en los antiguos homínidos conocidos.
En lo demás, los
rasgos son bastante primitivos.
- 60-45 kg. Este rango de variación indica un dimorfismo extremo.
- El cráneo MRD tiene una cresta sagital bien desarrollada y una cara robusta, larga y prognática. Capacidad craneal 365–370 cc, similar a S. tchadensis y más baja que la de cualquier individuo conocido de A. afarensis.
- Orificios auditivos pequeños y elípticos como en los grandes simios actuales.
- Esmaltes gruesos (1,5-2 cm). El desgaste de las coronas presenta fuerte inclinación, mientras que en el resto de los australopitecos es plano. La masticación se produce con movimientos más verticales de la mandíbula. Mandíbula algo menos robusta que en afarensis, con arcada dental estrecha de hileras paralelas. Sínfisis mandibular muy inclinada hacia atrás y dentición en forma de U, como en los chimpancés. Engrosamiento del esmalte, con una dentición anterior reducida. Paladar estrecho y poco profundo. Apertura nasal redondeada y ligeramente unida hacia el maxilar, mientras que en afarensis la apertura está más separada y presenta una prominente cresta lateral.
- Incisivos laterales inferiores relativamente más grandes que los de afarensis.
- Caninos grandes, asimétricos y plantados en raíces largas y robustas. Raíces de los caninos superiores implantadas verticalmente, lo que sugiere un rostro moderadamente inclinado hacia atrás. Caninos mandibulares estrechos y asimétricos. Raíz de canino más grande del registro homínido.
- Complejo canino/premolar: Morfología característica de los homínidos tempranos con corona canina baja, pero con un diente más afilado mesiodistalmente que en A. afarensis y con unas raíces de los caninos más grandes y probablemente dimórficas, lo que sugiere una diferencia en la función adaptativa canina entre A. anamensis y A. afarensis.
- Primer premolar inferior P3 con dos raíces bien separadas y una corona muy asimétrica, con una cúspide, como en los simios antropoides.
- Esta especie aporta la primera evidencia de incremento en el tamaño de los molares, que se asocia con el consumo de alimentos más duros y su vida en la sabana. Grandes molares. Molares superiores M1 y M2 de tamaño comparable.
- Tibia robusta con superficies articulares parecidas a las humanas. Marcha con la rodilla en extensión y tibia sometida al peso del cuerpo, con menor desgaste de la suspensión. En Kanapoi se recuperaron los extremos superior e inferior de una tibia derecha, que presentan indicios de bipedia: las epífisis tienen mayor cantidad de tejido esponjoso para absorber el mayor peso que un bípedo transmite por las articulaciones de la extremidad inferior; además, la orientación de la epífisis inferior es más similar a la de los humanos. Algunas plesiomorfias, podrían indicar una bipedación todavía incipiente.
- La articulación del codo, con una fosa olecraneana poco profunda, recuerda la humana. Los huesos de los brazos y manos sugieren adaptaciones a la vida arbórea.
- Dimorfismo sexual marcado, mayor que en afarensis, que supone un problema para la catalogación.
- Las vértebras recuperadas en Assa Issie indican bipedalidad habitual. En contraste con las líneas de evidencia craneodentales, algunos aspectos de la morfología vertebral en A. anamensis parecen más derivados que los de sus descendientes A. afarensis (Meyer y Williams, 2019).
 |
| Algunas diferencias del Australopithecus anamensis con el chimpancé y el ser humano moderno. |
 |
Arriba, vistas oclusales de izquierda a derecha, KNM-KP
29281, KNM-KP 29287 y KNM-KP 31713. En la parte inferior, algunas mandíbulas de
afarensis: LH 4, AL 123-23, AL
333w-60, AL 266-1, AL 400-1a, AL 277-1 y AL 198-1. Las flechas señalan la
inflexión anterolateral de la línea oclusal. En anamensis los caninos están situados anteriormente con respecto a
P3 mientras que en afarensis se
sitúan medialmente. La barra indica una escala de 0–4 cm. De C. V. Ward et al
(2010).
|
Hábitat
- Aralee Issie (ARI) y Mesgid Dora (MSD), en Woranso-Mille, estaban rodeados predominantemente de hábitats abiertos, como matorrales. El análisis de desgaste ha revelado que los ungulados consumían todos los tipos de dieta (hierbas, hojas, frutas) en parecidas proporciones. Los comportamientos locomotores de los bóvidos, deducidos a partir de los astrágalos y falanges recuperados, están asociados con todos los tipos de hábitats. En conjunto, estos resultados sugieren hábitats heterogéneos (mosaicos), con probables áreas de gran densidad de vegetación a lo largo de un paleorío y regiones más abiertas (bosques, pastizales) disponibles lejos del río (Sabrina C. Curran y Yohannes Haile-Selassie, 2016).
Discusión filogenética
Los Australopithecus se diferencian de los ardipitecinos fundamentalmente en:
· Los dientes poscaninos, de mayor tamaño y con el esmalte más grueso.
· Diferente forma del canino superior.
· Mayor dimorfismo.
· Locomoción, con mejores adaptaciones a la bipedación.
- Leakey et al (1995) aceptan la relación anamensis -> afarensis, mientras que ramidus sería una especie hermana. Ramidus y anamensis ocuparon distintos nichos adaptativos (White, 2009).
- White (2002) acepta la posibilidad de una relación ramidus -> anamensis, con un cambio anatómico relativamente rápido.
- anamensis y afarensis son dos cronoespecies (anagénesis)
- Ward et al (1999)
- Ward, Leakey y Walker (2001)
- Kimbel et al (2006) señalan una secuencia temporal anamensis de Kanapoi -> anamensis de Allia Bay -> afarensis de Laetoli -> afarensis de Hadar y consideran que la evolución homínida en este periodo no fue tan ramificada como se pensaba.
Leakey et al (1998) propusieron que
- O estos homínidos consituían una especie única que fue evolucionando.
- O que existieron tres especies (ramidus, anamensis y afarensis) con relaciones filogenéticas imprecisas.
- Skelton y McHenry (1992) advirtieron sobre la existencia de posibles homoplasias en la dentición de estos homínidos (rasgos similares en especies independientes desarrollados para hacer frente a presiones adaptativas similares).
Haile-Selassie et al (2019), consideran que BEL-VP-1/1 e incluso los dientes de Fejej (Ward et al, 2014) deberían clasificarse como A. afarensis por lo que esta especie habría coexistido en el tiempo con A. anamensis.
Adam van Arsdale: Australopithecus anamensis
Te puede interesar:
|
Excelente. Agradezco la suerte de éste humilde homínido; el hallazgo este magnífico blog me permite una notable documentación y un recurso para contrastar etc. Por supuesto, y con permiso, os enlazo en mi blog. Un millón de gracias y, por favor, continuad este atractivo trabajo.
ResponderEliminarAndrés Ruiz Segarra
(http://entrehominidos.blogspot.com.es/)
Gracias por tus palabras, Andrés. Por supuesto, todos los contenidos de este blog están a tu disposición y a la de quienes gusten, pues su objetivo es precisamente la difusión del conocimiento.
Eliminarmuchas gracias por compartir la información
ResponderEliminarGracias a ti por tu interés.
ResponderEliminar