¿Preneandertal?
|
¿Heidelbergensis?
|
|
MIS15-MIS13 621-478 ka
|
Mauer, Boxgrove
|
|
MIS12 478-424 ka
|
Arago
|
|
MIS11 424-374 ka
|
Sima de los Huesos
|
Ceprano, Aroeira
|
MIS10-MIS9 374-300 ka
|
Swanscombe
|
|
MIS8 300-243 ka
|
Atapuerca Galería
|
Bilzingsleben
|
MIS7 243-191 ka
|
Pontnewydd, Steinheim, Ehringsdorf, Tourville, Saccopastore, Petralona
|
En la Sima de los Huesos (Atapuerca, Ibeas de Juarros, Burgos, España) se hallaron más de 6.700 fósiles (80% de los conocidos del Pleistoceno Medio en todo el mundo) de unos 28 individuos: un niño, doce adolescentes, diez jóvenes, y diez adultos de más de 25 años de los que solo tres superarían los 35. Ninguno superaría los 45. Sexos equitativamente representados, con una diferencia de tamaño de un 10%. 0,4 Ma. Los primeros, descubiertos en 1976, por el equipo de Trino Torres. Lee J. Arnold et al (2014) han obtenido una datación mínima de 427 ± 12 ka.
El análisis del ADN nuclear ha confirmado la pertenencia de esta población al linaje neandertal (Meyer et al, 2016)
 |
| Estratigrafía, plano y sección de la Sima de los Huesos. Sala et al, 2016. |
 |
| Los cráneos de la Sima de los Huesos. Sala et al, 2016. |
 |
| Sima de los Huesos. Cráneo 17. Crédito de la foto: Javier Trueba / Madrid Scientific Films. |
 |
| Sima de los Huesos. Cráneo 15. Crédito de la foto: Javier Trueba / Madrid Scientific Films. |
 |
SCi: Sala de los Cíclopes;
OC: Chimenea obstruida.
C1, C2, C3: Chimeneas
SH: Sima de los Huesos.
SR: Rampa.
ST: Cima.
SRB, SRM, SRA: Pozos de sondeo en Rampa Baja, Rampa Media y Rampa Alta.
|
La muestra de Homo de la Sima de los Huesos presenta fracturas en una frecuencia muy alta. Los huesos largos muestran un patrón caracterizado por el dominio de las fracturas transversales del eje largo, circunferencias completas y bordes de la fractura con ángulos rectos y superficies irregulares. Nohemi Sala et al (2015) han analizado dichas fracturas. Según las conclusiones, la mayoría de ellas se produjo después del entierro debido a la presión de los sedimentos. Una pequeña proporción de fracturas podrían ser debidas a un traumatismo contundente producido por la caída libre de 13 m que constituye el acceso a la cámara de la Sima de los Huesos. El Cráneo 17, sin embargo, muestra dos fracturas perimortem con depresión en el hueso frontal, que se interpretan como dos episodios traumáticos. El tipo de lesión, su ubicación, la gran similitud de las fracturas en forma y tamaño, y las diferentes orientaciones y trayectorias implícitas de las dos fracturas sugieren que fueron producidos con el mismo objeto, por un individuo diestro, durante un conflicto interpersonal cara a cara. La presencia de múltiples golpes implica una intención de matar. Este hallazgo demuestra que la violencia interpersonal letal es un comportamiento humano antiguo y tiene implicaciones importantes para la acumulación de cuerpos en el lugar, apoyando a un origen antrópico (Nohemi Sala et al, 2015b). Nohemi Sala et al (2016) han analizado mediante tomografía computerizada los ocho cráneos que muestran traumas típicos perimortem. Además del Cráneo 17, la violencia interpersonal es la probable causa de traumas en los cráneos 5 y 11 y la más posible en los otros cinco cráneos.
 |
| Sima de los Huesos. Cráneo 6. Javier Trueba. |
David Rabadà i Vives (2015), por el contrario, ha revisado la literatura publicada llegando a la conclusión de que el yacimiento fue originado mediante diversos mecanismos tafonómicos, con una baja tasa de sedimentación y el aporte eventual de cadáveres.
- La competición entre Homo y depredadores por la cavidad pudo originar la muerte de los humanos cerca de la fosa. Esto podría explicar la distribución de la mortalidad, ya que la depredación es un accidente y los adultos jóvenes y los adolescentes sufren los accidentes en mayor medida. Los caninos y otros carroñeros actuaron más tarde.
- La ausencia de herbívoros podría deberse a que los rebaños poblaban las llanuras abiertas lejos de la Sima de los Huesos, con un ambiente boscoso y montañoso y los depredadores transportan los cadáveres solo unos metros.
- Los flujos de agua ocasionados por las tormentas, transportaron, dispersaron, mezclaron y produjeron la abrasión de los restos humanos y del hacha de mano.
- Estos procesos fueron acompañados de una baja tasa de sedimentación con una aportación eventual de cadáveres durante un largo periodo de tiempo.
- Cuando la cavidad fue cerrada por los desprendimientos quedando como principal entrada el pozo actual, los osos cayeron por él accidentalmente o murieron mientras hibernaban.
 |
| Sima de los Huesos. Cráneo 9. Crédito de la foto: Javier Trueba / Madrid Scientific Films. |
- 17 cráneos (J. L. Arsuaga et al, 2014). Son muy similares entre sí y distinguibles de los de otras poblaciones europeas de datación similar, lo que indica que estas poblaciones pertenecían a diferentes demos.
- El patrón morfológico es consistente con características neandertales derivadas en la cara anterior y la bóveda, muchas de las cuales están relacionadas con el aparato masticatorio incluyendo su uso como tercera mano, con gran desgaste de los incisivos. Por el contrario, las características endocraneales son sorprendentemente primitivas y no recuerdan los ragos neandertales.
- Los autores sitúan esta población en las raíces de la evolución neandertal, sugieriendo que la modificación facial fue el primer paso, dentro de un patrón de mosaico, con diferentes módulos anatómicos y funcionales evolucionando a un ritmo diferente.
- Están presentes características de los neandertales clásicos, como la forma triangular de la cara y los arcos superciliares, prognatismo, mandíbulas inferiores robustas, patrones de las cúspides dentales y dientes pequeños en la parte posterior de la mandíbula, pero no se hallan otras como el moño occipital o el desarrollo cerebral.
- Taxonómicamente, no es posible clasificar esta población dentro de heidelbergenesis, pero tampoco en neanderthalensis.
- Estos resultados y la comparación con otros rasgos craneales confirman que los neandertales evolucionaron en forma de mosaico. Los rasgos relacionados con la masticación (anatomía dental, facial y mandibular) condujeron al proceso de neanderalización, mientras que la anatomía neurocraneal debe haber adquirido una condición completamente neandertal considerablemente más tarde (Poza-Rey, Gómez-Robles y Arsuaga, 2019).
- 10 individuos padecían cribra orbitaria, extensión de perforaciones en el techo de la cavidad de las órbitas. En varios casos se observan traumatismos curados. La forma no es tan alargada como en los neandertales, sin la protuberancia occipital y el aplanamiento lamboidal. La anchura máxima se sitúa en la parte inferior del cráneo, en las crestas mastoideas, lo que proporciona un perfil con forma pentagonal baja (morfología primitiva; los neandertales tienen un contorno posterior redondeado y los sapiens un contorno pentagonal alto). En cuanto al hueso occipital, no presentan la proyección bilateral del torus occipital ni la depresión media de los neandertales. Por encima del torus occipital aparece un área deprimida que pudira ser un primer estadio de la fosa suprainiánica de los neandertales. El borde superior de la escama temporal es alto como en neandertales y sapiens (en erectus es bajo). Apertura nasal amplia pero sin presentar el borde inferior afilado de los neandertales. Apófisis mastoideas bien desarrolladas y proyectadas hacia abajo (en los neandertales poco proyectadas). En resumen, Arsuaga et al (1997) enfatizan la presencia de características arcaicas ausentes en los neandertales junto con características derivadas pero menos desarrolladas que en los neandertales y el parecido con los cráneos de Saccopastore.
- Menor tamaño que neandertales y humanos modernos (también menor grado de encefalización):
- Cráneo 4: 1.390 cc.
- Cráneo 5: 1.125 cc.
- Cráneo 6: 1.220 cc.
- Los Homo de la Sima de los Huesos no difieren de los neandertales en ninguna de las variables relacionadas con los tamaños absolutos y relativos y la forma de los canales semicirculares, incluyendo un relativamente pequeño canal posterior y un canal lateral relativamente grande. En contraste, no está presente la baja colocación del canal posterior, característica distintiva de los neandertales. El índice de forma coclear, es bajo, lo que puede ser una característica derivada distintiva de esta demo. Rolf Quam et al, 2015.
- De acuerdo con el análisis dental, todos los individuos de la SH eran diestros. En cuanto a los endocastos, la proporción de diestros y no diestros en la SH (4/6) difiere en gran medida de la proporción de diestros/zurdos en las poblaciones humanas modernas (9/1). Esto representa un menor grado de lateralización del cerebro en el grupo SH o por lo menos un grado diferente de lateralización que en los grupos humanos modernos (Eva María Poza-Rey, Marina Lozano y Juan Luis Arsuaga, 2015).
- 20 mandíbulas (medio centenar de piezas), cuatro de ellas enteras. Son robustas, sin mentón. Cuerpo mandibular más bien bajo, con el borde inferior ancho, plano, paralelo a la fila de dientes y con un acusado reborde. Ramas ascendentes largas, poco altas y con el borde superior poco escotado. En seis individuos se observa una artrosis degenerativa de la articulación del hueso temporal y la mandíbula.
- Más de 500 dientes.
- Incisivos superiores en pala.
- Caninos con cíngulos gruesos.
- Premolares no molariformes.
- Molares decrecientes, ya del primero al segundo, tanto en largo (igual que los neandertales) como en ancho (típicamente moderno).
- Son frecuentes las hipoplasias en el esmalte (siete individuos), pero con una proporción baja en el total, lo que indica un menor estrés alimenticio que el que se ha advertido en otros grupos humanos. Las hipoplasias se producen a los tres años, edad del destete.
- No se ven signos de caries, que aparecerían con el consumo de cereales, pero sí de la utilización de mondadientes. Hay marcas en la superficie externa de los dientes anteriores causadas por filos silíceos al utilizar la dentadura como herramienta.
- Elevado desgaste dental, incluso en los dientes de individuos jóvenes.
- En los dientes posteriores, aparecen estrías con orientaciones diversas y agujeros en las superficies oclusales, lo que indica una dieta muy abrasiva.
- Las estrías verticales indican una dieta cárnica.
- Las estrías en otras direcciones indican una dieta vegetal, más difícil de masticar, que exige movimientos en horizontal de las mandíbulas.
- Los dientes anteriores están muy desgastados lo que indica su uso como herramienta.
- Para María Martinón-Torres y José María Bermúdez de Castro (2011) la morfología de estos dientes es completamente neandertal y por ello estos individuos de Atapuerca pertenecen a un grupo o especie diferenciados, pues estos rasgos están ausentes en los restos de los demás yacimientos de similar datación.
- El M1 superior presenta un mosaico de características neandertales y sapiens (María Martinón-Torres et al, 2013).
- El M1 inferior se parece al de los neandertales (María Martínez de Pinillos et al, 2015).
- García-Campos et al (2020) han evaluado el tejido dental de los caninos permanentes para caracterizar el patrón y el grado de dimorfismo sexual. La proporción de sexos observada es 5: 9 (N m : N f) En términos generales, las muestras dentales de Sima de los Huesos y Krapina tienen un grado de dimorfismo sexual que no supera la de los humanos modernos.
- Los molares exhiben en promedio un esmalte absoluto y relativo grueso en estimaciones 2D y 3D, tanto en la corona completa como en el esmalte lateral. Esta condición primitiva se comparte con la mayoría de los hominini extintos y sapiens, a excepción de los neandertales y algunas muestras aisladas. Por el contrario, los mapas de distribución del esmalte revelan un patrón de distribución similar al de los neandertales, con mayor gruesa en las cúspides linguales y más distribuido periféricamente, en comparación con H. antecessor y humanos modernos. Debido a la posición filogenética de la población de la Sima de los Huesos, la condición gruesa en los molares podría representar la persistencia de la condición plesiomórfica (Martín-Francés et al, 2020).
- 42 restos de clavículas, 114 de escápulas, 645 restos de vértebras, 466 de costillas que manifiestan un cuerpo ancho, una plesiomorfia heredada de los Hominini tempranos (Arsuaga et al, 2015).
- Las clavículas son más largas que las de los humanos modernos y muestran una curvatura de tipo II en el plano coronal que está presente en todos los Homo anteriores al sapiens. Este carácter se ha relacionado con una posición más lateral y superior de las escápulas.
- Conducto medular largo en los atlas y corto en los axis, que son anchos en comparación con los modernos.
- La ausencia de costillas torácicas medias completas, hace que sea difícil evaluar la forma del esqueleto costal, pero posiblemente fuera más amplio: el tamaño dorsoventral de la única primera costilla recuperada, es mayor que en los humanos modernos y los neandertales y una segunda costilla incompleta sugiere que era mas larga dorsoventralmente que la de Kebara 2.
- La pelvis es muy diferente a la de los humanos modernos, muy amplia y elíptica, con un sacro muy ancho, pronunciada abertura de las alas ilíacas en sentido lateral, como en los australopitecinos y ramas púbicas largas. También es más amplia que la de los neandertales.
- Reducida lordosis lumbar, una característica derivada compartida con los neandertales.
- 110 restos de húmeros, 65 de cúbitos y 89 de radios (49 de adultos).
- Laura Rodríguez García, en su tesis doctoral sobre los huesos largos (2013), considera que estos Homo eran igual de eficientes que los HAM. Incluso la flexión y giro del antebrazo serían más eficientes. Los huesos de las extremidades inferiores poseen más cortical, lo que les permitiría soportar mucho más peso.
- Laura Rodríguez et al (2015) analizaron las propiedades de la sección transversal de la muestra radial en los puntos medios del eje y del cuello sin encontrar diferencias entre los Homo de la Sima de los Huesos, los neandertales y los humanos recientes. Sin embargo, debido a la mayor longitud del cuello del radio, la demo de la Sima de los Huesos disfrutaba de palanca y movimientos de pronación-supinación y flexión-extensión más eficientes que los de los neandertales o los humanos recientes.
- En el húmero, la cabeza articular es oval con un ángulo de torsión bajo. Varios refuerzos en la zona articular del codo.
- La cavidad glenoidea es más alta y estrecha que en los humanos modernos, lo que se refleja en un bajo índice glenoideo. Es dominante la posición dorsal del surco axilar del músculo redondo menor, como en los neandertales. La curvatura de las clavículas en el plano transverso cae dentro de la variación normal en los humanos modernos; sin embargo, en el plano coronal, es similar a la de los neandertales. La mayoría de los húmeros y radios son similares a los de los neandertales. La mano muestra características propias del agarre de precisión, similares a las descritas para neandertales y humanos modernos. Como en los neandertales, el poderoso agarre de precisión se ve reforzado por un pulgar robusto y una musculatura flexora bien desarrollada (Arsuaga et al, 2015).
- 173 restos de fémures, 20 de rótulas, 89 de tibias y 105 de peronés. Los huesos largos son muy fuertes.
- Grandes cabezas femorales en algunos individuos, un rasgo que apareció por primera vez durante el Pleistoceno Medio en África y en Europa. El peso medio, a partir de las cabezas femorales de cinco adultos, ha resultado 69,1 kg, 6,3 kg por debajo del peso medio de los neandertales. Los fémures muestran el patrón morfológico plesiomórfico que aparece en los miembros más tempranos del género Homo mientras que las tibias son similares a las de los neandertales. (Arsuaga et al, 2015).
- 121 huesos del carpo, 103 del tarso, 93 metacarpianos, 80 metatarsianos, 208 falanges de pies y 304 falanges de manos. En los restos de niños, se dan varios casos de líneas de Harris, consecuencia típica de enfermedades infantiles.
- En la campaña de 2012 se halló una falange distal del meñique del pie de un niño. El hallazgo de un hueso tan pequeño del pie apoya la hipótesis de que la Sima de los Huesos era un lugar de enterramiento y descarta que los restos fuesen acumulados por depredadores.
- En comparación con los humanos modernos, el astrágalo de los neandertales muestra facetas maleolares laterales y cabezas talares amplias. En los Homo de la Sima de los Huesos, la faceta peronea es significativamente más amplia y la cabeza talar más estrecha; al igual que en los neandertales la tróclea es relativamente amplia con bordes paralelos mientras que los humanos modernos es relativamente estrecha y en forma de cuña. Todos los Homo del Pleistoceno (no sapiens) muestran unos astrágalos con una tróclea más alta que en los humanos modernos, permitiendo una gran capacidad de flexión dorsal y plantar del tobillo. Los calcáneos de los neandertales son anchos, con una plataforma del astrágalo proyectada y un tubérculo largo. En los Homo de la Sima de los Huesos, los calcáneos son también anchos y la plataforma del astrágalo aparece incluso más proyectada. Los metatarsos de los humanos modernos, neandertales y Homo de la Sima de los Huesos son muy similares, excepto por la amplia base de los MTIII–V, una característica potencialmente derivada compartida por los Homo de la Sima de los Huesos y los neandertales. Las falanges proximales presentan hipertrofia del eje y la falange distal del pulgar muestra una tuberosidad distal expandida, como en los neandertales. (Arsuaga et al, 2015).
- Hioides y huesecillos del oído. I. Martínez ha deducido de su análisis la capacidad de habla de aquellos individuos.
- Pelvis 1 y columna lumbar (Elvis). 0,5 Ma. Bonmatí et al, 2018. Pelvis con fuertes inserciones musculares correspondientes a una piernas muy robustas. La morfología representa el patrón primitivo dentro del género Homo, con dimorfismo sexual en la forma del canal pélvico. La pelvis más ancha y mejor conservada de todo el registro fósil y la evidencia más antigua de un individuo de edad avanzada. Correspondió a un varón muy corpulento que padeció importantes enfermedades degenerativas (signos de deformidad cifótica lumbar, espondilolistesis y enfermedad de Baastrup) desde mucho antes de morir, con más de 45 años de edad. Debió medir entre 1,70-1,75 m y pesar 90-95 kg. Dichas enfermedades debieron de producirle intensos dolores en las zonas lumbar y pélvica que le obligaban a caminar encorvado y, probablemente, a utilizar un bastón para mantenerse erguido. Debido a sus mermadas condiciones físicas, este individuo no estaba en condiciones de cazar ni de llevar a cabo otras tareas útiles para su comunidad, lo cual ha llevado a la conclusión de que, además de alimentación, el anciano debió también de recibir ayuda para seguir el ritmo de la marcha de su gente, un grupo de cazadores nómadas que se desplazaban junto a las manadas de herbívoros de los que se alimentaban.
- Agamenón, Atapuerca 4. Caja craneal completa, sin su parte facial. Se observa una artritis avanzada en los cónditos occipitales, con importante dificultad y dolor en los movimientos de la cabeza. El frontal huido, la bóveda baja y el occipital alargado y anguloso hacia atrás del cráneo 5 no son compartidos en el cráneo 4. La bóveda relativamente baja y el frontal estrecho son rasgos comunes.
- Miguelón, Atapuerca 5 (AT 700). Descubierto en la Sima de los Huesos (Atapuerca) entre 1992 y 1993 por Juan Luis Arsuaga. Su antigüedad se estima en 0,3 Ma; su capacidad craneal es de 1.125 c.c. Se trata del cráneo más completo de heilderbengensis descubierto hasta ahora. (Arsuaga et al. 1993; Johanson and Edgar 1996). Se ha identificado la mandíbula que le corresponde.
- Morfología facial similar a la de los neandertales (prognatismo, huesos nasales sobresalientes y amplios, posición avanzada de la dentición inferior, espacio retromolar). En relación al neurocráneo, la cara es muy grande. Placa infraorbitaria ligeramente cóncava, con el margen inferior ligeramente curvado (en los neandertales la placa infraorbitaria es plana con el margen recto).
- Macizo maxilar grande, proyectado hacia delante sin fosa canina entre el saliente nasal y el pómulo ni fosa maxilomalar entre el pómulo y la pared de las raíces de los molares superiores. Orbitas grandes y casi redondas. Prognatismo y adelantamiento del arco de los dientes que deja un espacio retromolar entre el talón de la última muela inferior y el borde anterior de la rama ascendente de la mandíbula.
- Padecía varias alteraciones en la boca. La más grave, una infección en el lado izquierdo tras la fractura de un diente y aplastamiento del maxilar debido a un golpe con una piedra.
- Del estudio de la lateralidad del cerebro, se concluye que era diestro.
- Benjamina. Individuo de 10 años de edad. Padeció craneosinostosis, enfermedad que produce una sutura anticipada de los huesos craneales, lo que se traduce en un retraso sicomotor.
- AT 624. Individuo infantil con un fuerte golpe en el lateral del torus supraorbitario izquierdo, con signos de regeneración ósea.
Entre 2015-2018, restos craneales de una hembra de unos 13 años de edad (Sara).
 |
| Cráneo número 5 hallado en la Sima de los Huesos |
La estatura media se ha estimado a partir de 24 huesos largos completos de las extremidades superiores e inferiores en 163,6 cm, 3 cm más que la de los neandertales. Esta estatura es similar a la de los humanos modernos de latitudes medias y apareció por vez primera hace 1,6-1,5 Ma. El nivel de dimorfismo es similar al de los humanos modernos (Arsuaga et al, 2015). Fémur, tibia y peroné presentan una enorme robustez, en la línea de neandertales, ergaster, erectus y otros Homo arcaicos.
Paredes óseas más gruesas y diámetros exteriores mayores implican mayor resistencia a las fuerzas de combadura y torsión, pero también a la carga axial (vertical), que es la que está provocada básicamente por el peso corporal del individuo. Además, el fémur presenta secciones de forma muy circular, mientras que en los humanos actuales suele ser más bien elíptica o en forma de almendra, lo que indica patrones de fuerzas diferentes (Laura Rodríguez et al, 2018). Los huesos largos tienen, en relación con su tamaño, un mayor volumen óseo (volumen de tejido óseo sobre el volumen total del hueso) que los huesos equivalentes en una nuestra muestra de humanos recientes. Los Homo de la Sima de los Huesos tenían en promedio huesos más pesados que los humanos actuales del mismo tamaño. A partir del peso femoral se puede estimar que el peso total del esqueleto era un 36% mayor que en el de los humanos recientes de tamaño corporal equivalente (Carretero et al, 2018).
En 1998 apareció un bello bifaz (Excalibur) de perfecta talla y atractivo color marrón y rojo. Se ha interpretado en términos simbólicos, pues nunca fue utilizado. Datado en 0,4 Ma. En la campaña de 2012 se encontró un bifaz adicional.
Paredes óseas más gruesas y diámetros exteriores mayores implican mayor resistencia a las fuerzas de combadura y torsión, pero también a la carga axial (vertical), que es la que está provocada básicamente por el peso corporal del individuo. Además, el fémur presenta secciones de forma muy circular, mientras que en los humanos actuales suele ser más bien elíptica o en forma de almendra, lo que indica patrones de fuerzas diferentes (Laura Rodríguez et al, 2018). Los huesos largos tienen, en relación con su tamaño, un mayor volumen óseo (volumen de tejido óseo sobre el volumen total del hueso) que los huesos equivalentes en una nuestra muestra de humanos recientes. Los Homo de la Sima de los Huesos tenían en promedio huesos más pesados que los humanos actuales del mismo tamaño. A partir del peso femoral se puede estimar que el peso total del esqueleto era un 36% mayor que en el de los humanos recientes de tamaño corporal equivalente (Carretero et al, 2018).
Aunque los Homo de la Sima de los Huesos estaban menos encefalizados que los neandertales, comparten con estos muchas características poscraneales. La mayoría de estas características parecen ser retenciones plesiomórficas o de polaridad filogenética incierta, pero otras son consideradas apomorfias neandertales. Sin embargo, el conjunto completo de características derivadas presentes en los neandertales aún no se observa en la población de la Sima de los Huesos (Arsuaga et al, 2015).
- Algunas especializaciones neandertales no están presentes en los Homo de la Sima de los Huesos, tales como la orientación lateral de las apófisis transversas lumbares, la articulación carpometacarpiana del pulgar menos en forma de silla una rama superior del pubis extremadamente delgada.
- Algunos rasgos relevantes, son polimórficos en el Homo de la Sima de los Huesos, como la morfología general del radio, la morfología de la frontera axilar de la escápula y la forma de la porción distal del húmero. En los neandertales se produce una pérdida de variación y fijación de caracteres.
En 1998 apareció un bello bifaz (Excalibur) de perfecta talla y atractivo color marrón y rojo. Se ha interpretado en términos simbólicos, pues nunca fue utilizado. Datado en 0,4 Ma. En la campaña de 2012 se encontró un bifaz adicional.
Otros hallazgos en España
Atapuerca-Galería, en la comunicación con la Sala de los Zarpazos. 0,3 Ma.
- AT 76-T1H. Fragmento de mandíbula derecha con dos molares hallado por Trinidad Torres (1976). La presencia de espacio retromolar, el toro mandibular y los tamaños de los últimos molares jutifican su asignación al demo de los preneandertales europeos.
- Fragmento de cráneo hallado en 1995 que comprende la parte posteromesial del parietal derecho, parte posterior de la sutura sagital, otro fragmento menor del parietal izquierdo y el tramo central de la sutura labdática, con una porción muy pequeña adherida del occipital que incluye el lambda. El parietal es muy grueso y fue hundido por un golpe que luego sanó.
- Restos líticos Modo 2.
- Del análisis polínico se desprende un periodo relativamente frío.
- En el nivel TD10 apareció una tecnología Modo 2 asociada a un procesamiento sistemático y direccional de carcasas, incluyendo eventos de caza, datado en 0,45 Ma. TD10-1 puede representar la evolución local desde el Modo 2 al Modo 3 (Andréu Ollé et al, 2013).
Pinilla del Valle (Madrid): 2 dientes.
Cueva del Congosto (Alcorlo, Guadalajara, España). Un metatarsiano y una vértebra cervical. Asociados a restos de animales con trazas de haber sido un basurero de hienas.
Sima de la Cueva del Ángel (Lucena, Córdoba, España). Un fémur. Los trabajos que se desarrollaron en la Cueva del Ángel, en los años 1995, 1996, 2002 y 2003, permitieron detectar la existencia de un potente depósito arqueológico con una gran amplitud cronológica que se situaría entre los 0,48-0,1 ka. La estratigrafía que es visible en más de seis metros de potencia, no parece mostrar periodos de interrupción del hábitat humano, muy al contrario, la superposición de hogares en la misma -que pueden llegar a alcanzar hasta los 60 cm. de espesor- confirman que las perduraciones son muy importantes. Podría constituir uno de los últimos lugares de ocupación del heidelbergenesis, en un momento en que los neandertales poblaban Europa.
Cueva de Lezetxiki (Mondragón, Guipúzcoa, España). Húmero completo de un individuo femenino. Datación incierta. En 2014 fue datado directamente en un mínimo de 165 ka.
Tossal de la Font de Vilafamés (Castellón, España). Fragmento distal de húmero y fragmento de coxal.
Bolomor (Valencia, España). Varios dientes y fragmento de peroné.
Valdegoba (Burgos, España). Mandíbula, dientes y tres restos poscraneales.
Un diente de infante, hallado en la Cueva de Mollet, Parc de les Coves Prehistòriquess de Serinyà (Pla de l’Estany), fue datado en 215 ka.
Un diente de infante, hallado en la Cueva de Mollet, Parc de les Coves Prehistòriquess de Serinyà (Pla de l’Estany), fue datado en 215 ka.
En febrero de 2013, se publicó la noticia del hallazgo en una cueva de Conca de Dalta (Lérida, España) de restos neandertales y herramientas achelenses, datados en 200 ka.
 |
| Hallazgos de la Sima de los Huesos. Crédito: Javier Trueba Madrid Scientific Films. |
 |
| Saccopastore 1. Foto: A. Mounier. |
Hallazgos en Italia
En Saccopastore, cerca de Roma, aparecieron entre 1929-1935, un cráneo humano masculino y otro femenino, de características arcaicas, similares a los cráneos de la Sima de los Huesos (Arsuaga el al, 1997).
- Saccopastore 1 está casi completo y muestra arcos superciliares muy grandes, frente inclinada baja y bóveda occipital redondeada. Los inicios del tercio medio facial están amplidos, con grandes dientes y nariz.
Según una reciente revisión estratigráfica de Marra et al (2015) los cráneos se depositaron hace ca 250 ka, en el inicio de MIS 7.
 |
| Reproducción del cráneo de Steinheim |
Hallazgos en Alemania
- Steinheim (Stuttgart, Alemania). 1934. Cráneo casi completo, deformado y restaurado. Unos 1.100 cc. Largo y estrecho. Moño occipital y apomorfias de neandertal poco avanzadas, pero la cara no muestra rasgos neandertales. 0,225 Ma (Adam, 1954, 1985). Por su pequeño tamaño, posiblemente corresponda a una hembra. Caja craneana bastante esférica. Órbitas relativamente pequeñas, coronadas por unos prominentes rebordes supraorbitarios en doble arco. Orificio nasal bajo y ancho. Pómulos retraídos formando una pequeña fosa y no engrosados como en los neandertales. Fosa canina.
- Ehringsdorf (Alemania) Restos craneales y poscraneales de varios individuos. Rasgos de neandertal arcaico. 0,25-0,2 mda.
- Reilingen (Alemania). Cráneo incompleto. Algunos rasgos afines a neandertal.
Hallazgos en Reino Unido
- Swanscombe (East Anglia, Reino Unido). Occipital muy completo (1935), parietal izquierdo (1936) y derecho (1955) de hembra joven adulta. 1.300 cc. Paredes gruesas. Occipital redondeado. Ambas características lo asemejan a los neandertales, pero la opinión no es unánime. 0,3-0,4 Ma (Stringer y Hublin, 1999). Gran número de útiles líticos achelenses, incluyendo bifaces, raspadores y denticulados.
- Hoxne, Clacton-on-Sea, Barnham e High Lodge (East Anglia, Reino Unido). Registro fósil y de industria Modo 2. Cultura clactoniense.
- En Clacton-on-Sea se recuperó en 1911 una punta de lanza en madera de tejo, que mide 38,7 cm de largo y 39 mm de diámetro, datada en 420 ka. Representa el artefacto de madera más antiguo conocido.
- En Barnham, Suffolk, se identificó un cambio tecnológico, desde núcleos simples y escamas a hachas de mano. Los dos conjuntos deben corresponder a poblaciones con distinto origen (Ashton et al, 2017).
- Pontnewydd (Norte de Gales). Compton y Stringer, 2015. Fragmentos de maxilar y tibia. 17 dientes. Hallados entre 1980-1995. 0,251-0,195 Ma. Asociados con hachas de mano y artefactos levallois.
Hallazgos en Francia
- Biache-Saint Vaast. tres fragmentos de cráneo atribuidos al mismo individuo. Se observan rasgos de los neandertales clásicos. Trazado de la red meníngea avanzado, parecido con Swanscombe y con neandertal.
- Biache 1. Asociado a abundante industria musteriense tipo La Ferrasie. Solo se preserva la mitad posterior, que presenta morfología neandertal.
- Biache 2, fragmentado y mezclado con fauna. Muy robusto.
- Tourville-la-Rivière. Tres huesos largos de la misma extremidad superior izquierda de un joven o adulto, descubiertos en 2010 por A. Cottard y A. Thomannde en la capa D2inf del yacimiento al aire libre de Tourville-la-Rivière en el valle del Sena en el norte de Francia, donde reposaron tras un transporte fluvial postdeposicional. Las dataciones de hueso humano y dientes de animales asociados producen un rango de edad para el sitio de 226-183 ka (finales de MIS 7). Se recuperaron 726 elementos líticos asociados; la baja concentración, con excepción de una pequeña zona de talla, indica una ocupación efímera, probablemente relacionada con actividades de carnicería. Los hallazgos faunísticos están dominados por herbívoros de paisajes boscosos y no árticos y un clima templado interglacial; algunos restos muestran indicios de procesamiento humano. Jean-Philippe Faivre et al (2014) atribuyen los restos humanos a Homo neanderthalensis, aunque de forma no segura.
 |
| Petralona 1. Foto: A. Mounier. |
Hallazgos en Grecia
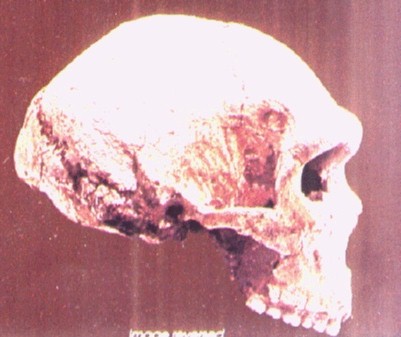
Petralona 1. Cráneo casi completo hallado casualmente en 1959, literalmente pegado a la pared de la cueva y cubierto por una capa de calcita formada quizá durante miles de años. El resto del esqueleto había desaparecido. El cráneo estaba a poco más de 20 centímetros del suelo de la cueva, por lo que muy posiblemente su dueño murió tumbado, con la cabeza apoyada en la pared. El arqueólogo y antropólogo griego Aris Poulianos bautizó el cráneo con el nombre de Archanthropus europeaus petraloniensis. Apomorfias neandertales en órbitas, rostro hacia delante, frente y bóveda craneanas hacia atrás con una cara alargada y plana y una mandíbula muy ancha. Su capacidad craneana es de 1.220 c.c., alta para ser erectus, pero baja para ser sapiens. Su edad se estima en 0,25-0,15 Ma (Grun, 1996). Robusto. Grandes rebordes supraorbitales en forma de doble arco y grandes órbitas oculares de forma aproximadamente cuadrangular. Pómulos hinchados, maxilar inflado con un margen inferior que desciende recto hacia el borde alveolar, como en el neanderthalensis. Orificio nasal muy ancho. Parte posterior angulosa y con gruesos rebordes transversales. Occipital primitivo.
Marathousa (Grecia). Artefactos líticos asociados con restos de elefante con marcas de corte. Datado entre 0,6-0,3 Ma.
{kind=link}
Marathousa (Grecia). Artefactos líticos asociados con restos de elefante con marcas de corte. Datado entre 0,6-0,3 Ma.
Adrián Pablos: Deportistas de hace 500 ka.
Conferencia de Marina Lozano. 2013.
Bibliografía:
- Kokkoros, P., and A. Kanellis. "Découverte d’un crane d’homme paléolithique dans péninsule Chalcidique." Anthropologie 64 (1960): 132-147.
- Stringer, Christopher B., F. Clark Howell, and John K. Melentis. "The significance of the fossil hominid skull from Petralona, Greece." Journal.of.Archaeological.Science 6.3 (1979): 235-253.
- Hennig, G. J., et al. "ESR7dating of the fossil hominid cranium from Petralona Cave, Greece." (1981): 533-536.
- Poulianos, Aris N. "Petralona cave dating controversy." (1982): 280-280.
- Rightmire, G. Philip. "Patterns of hominid evolution and dispersal in the Middle Pleistocene." Quaternary.International 75.1 (2001): 77-84.
- Beck, Curt W. "Archaeometric Clearinghouse."Journal of Field Archaeology 9.2 (1982): 279-280.
Sierra de Atapuerca
No hay comentarios:
Publicar un comentario