 |
| Datación de los principales yacimientos de Eurasia correspondientes al Paleolítico Inferior y eventos climáticos. |
Pocas respuestas
Los hallazgos de Dmanisi (Homo georgicus) marcan el inicio de la ocupación humana de Europa. Las evidencias más antiguas de esta ocupación están representadas por:
- Kozarnika, Bulgaria (1,6-1,4 Ma. Sirakov et al, 2010). Molar humano, industria de núcleo y lascas no achelense y huesos incisos que pueden ser el resultado de un comportamiento simbólico.
- Pirro Nord, Italia (1,6-1,3 Ma. Arzarello y Peretto, 2010). Hallazgos líticos.
- Atapuerca y Orce (España).
La presencia humana parece limitada al sur de la latitud 45ºN, constituida por grupos con tecnologías Modo 1 (Jöris, 2013).
 |
| Yacimientos del Paleolítico Inferior en Eurasia Occidental y Mediterráneo. Los círculos completos indican yacimientos líticos y los semicírculos yacimientos líticos y con restos humanos. |
La especie ñ
Los primeros fósiles europeos se datan entre 1,4 y 0,9 Ma. No ha sido identificada la especie a la que pertenecen. El equipo de Atapuerca considera que se trata de un nuevo taxón al que denominan provisionalmente "especie ñ", debido a que la mayor parte de los hallazgos corresponden al territorio español.
HOMO EN EUROPA ENTRE 1,3 Y 0,73 MA
| ||
PERIODO MA
|
FÓSILES
|
TECNOLOGÍA
|
1,4-0,9
|
No ha sido identificada la especie, que se denomina, provisionalmente, especie ñ
|
Modo 1 muy pobre
|
0,9-0,73
|
Homo antecessor
|
Modo 1 evolucionado
|
0,7-0,615
|
Homo antecessor en proceso de extinción.
Oleadas migratorias sin éxito demográfico.
|
Modo 1
Modo 2 pobre
|
0,615-0,2
|
Grado Heidelbergensis
|
Modo 2 pleno
|
0,2-0,027
|
Homo neanderthalensis
|
Modo 2
Modo 3. Musteriense.
|
 |
| Estratigrafía de los yacimientos ibéricos anteriores al subcrón Jaramillo. Joan García et al, 2014. |
Hombre de Orce
Descubierto en 1982 en las excavaciones dirigidas por J.
Gibert Clols en la cuenca de Baza (Venta Micena, Orce, Granada). Durante el
Pleistoceno Inferior existió allí un gran lago interior que desapareció hace 100 ka. No había cuevas y la vida de los homínidos se desarrollaba al aire libre. Gracias a las aguas termales sublacustres, a unos 36 ºC, el lago mantuvo un nivel freático relativamente permanente, con temperaturas más cálidas durante todo el año. Esto dio como resultado un alto nivel de productividad orgánica, especialmente en el Calabriense, lo que permitió el desarrollo de una comunidad de mamíferos rica y bien diversificada, similar a la de las sabanas africanas modernas. La alta biomasa de ungulados favoreció a los carroñeros, como las hienas y los homínidos, que dispusieron de abundantes canales, lo que explica las altas densidades de restos óseos conservados en los sedimentos distribuidos a lo largo de los alrededores del lago (José Manuel García-Aguilar et al, 2014). El análisis de microvertebrados de las unidades D1 y D2 de Barranco León revela que las temperaturas eran significativamente más altas que 13ºC, con condiciones húmedas prevalentes (Agustí et al, 2015).
El triángulo comprendido entre Orce y sus dos pedanías, Fuente Nueva y Venta Micena ofrece la mayor densidad de huesos fósiles de mamíferos cuaternarios conocida hasta el momento en Eurasia. La ausencia de suidos en las capas asociadas a restos humanos permiten datarlas entre 1,8-1,2 Ma (Bienvenido Martínez-Navarro et al, 2015).
En Fuente Nueva 3 y Barranco León 5, aparecieron restos modo 1, datados en 1,4-1,2 Ma. En Fuente Nueva hay evidencias de acción humana sobre herbívoros para consumir carne y tuétano.
El triángulo comprendido entre Orce y sus dos pedanías, Fuente Nueva y Venta Micena ofrece la mayor densidad de huesos fósiles de mamíferos cuaternarios conocida hasta el momento en Eurasia. La ausencia de suidos en las capas asociadas a restos humanos permiten datarlas entre 1,8-1,2 Ma (Bienvenido Martínez-Navarro et al, 2015).
En Fuente Nueva 3 y Barranco León 5, aparecieron restos modo 1, datados en 1,4-1,2 Ma. En Fuente Nueva hay evidencias de acción humana sobre herbívoros para consumir carne y tuétano.
- En Venta Micena:
- VM-0, fragmento parietal-occipital, presumiblemente de un niño. Se trata de la parte posterior de un cráneo de un niño de no más de cinco años, datado en 1,3-1,2 Ma. Al ser un resto tan fragmentario y presentar una cresa prominente, ha generado mucha controversia. Para Martínez-Navarro (2002) corresponde a una hembra de bóvido o cérvido. En 2003 se halló en Francolí (Tarragona) el esqueleto de un niño, en una necrópolis del siglo IV, con la misma cresta (Campillo et al, 2006), aunque Martínez-Navarro (2008) afirma que las morfologías de la sutura no se corresponden. Según el análisis inmunológico efectuado por Concepción Borja et al (1997), el fragmento corresponde a un homínido.
- VM1960, diáfasis humeral juvenil
- VM3961, parte intermedia de un húmero adulto.
 |
| Barranco León. Ubicación, estratigrafía y dataciones. Agustí et al, 2015. |
- En Barranco León D:
- BL5-0, parte de un molar, corona y raíz.
- Toro-Moyano et al (2013) sugieren que corresponde a Hippopotamus.
- Francesc Ribot et al (2015) han estudiado la posición estratigráfica y espacial de BL5-0 y su morfología, llegando a la conclusión de que perteneció a un humano (parte mesial de un dm2 izquierdo), confirmando la opinión de Arribas y Palmqvist (2002).
 |
| Venta Micena. VM-0. |
Hallazgos posteriores en Orce.
En Barranco León aparecieron fragmentos molares. Isidro Toro-Moyano et al, 2013, han descrito y datado el diente deciduo dm1 izquierdo de un homínido de unos diez años BL02-J54-100, Once boy, hallado en julio de 2002, asociado a 1.244 piezas líticas modo 1 y rectos óseos con fracturas y marcas de corte. La datación, de 1,4 Ma, convierte a este fósil en el resto homínido más antiguo de Europa Occidental. En las excavaciones de 2014 se descubrió un taller lítico.
- Francesc Ribot et al (2015) consideran que BL5-0 y BL02-J54-100 posiblemente podrían ser contiguos, correspondiendo con la desarticulación de un humano fallecido.
 |
| BL02-J54-100. El resto homínido más antiguo de Europa Occidental. Jordi Mestre. IPHES. |
- María Patrocinio Espigares et al (2013) han investigado los restos de una carcasa de elefante hembra (Mammuthus meridionalis) en Fuente Nueva-3 cuya extracción finalizó en 2003. La carcasa apareció incompleta (le faltan las extremidades y el cráneo), rodeada de 17 lascas de sílex y 34 coprolitos de hiena gigante caricorta (Pachycrocuta brevirostris), lo que sugiere un comportamiento competitivo entre los homínidos y esta especie. Los coprolitos aparecieron en los espacios que hubieran correspondido a las extremidades y los autores interpretan que los homínidos llegaros los primeros al cadáver, cortaron y se llevaron las patas y el cráneo, marchándose lo más rápido que pudieron. Después las hienas acabaron con el resto del cadáver.
 |
| Yacimientos de Atapuerca |
 |
| Sima del Elefante. Estratigrafía. |
Sima del Elefante
En la cueva de la Sima del Elefante, Sierra de Atapuerca, España, se han hallado los siguientes fósiles humanos:- ATE 9-1, en 2007, un fragmento de mandíbula datado en 1,2-1,1 Ma.
- La mandíbula no pudo ser atribuida a ninguna de las especies hasta entonces descritas. Incluye la región de la sínfisis, donde convergen las dos ramas mandibulares, la derecha conservada hasta el alveolo del primer molar y la izquierda hasta los alveolos del primer y segundo molar.
- La parte de la mandíbula es primitiva.
- La parte trasera es derivada.
- El desgaste dental es muy severo. Para compensarlo, los dientes sobre erupcionaron y descalzaron.
- Acumulaciones de cemento en las raíces dentales (hipercementosis).
- Presencia de infecciones.
- Dientes hacia fuera.
- Utilización de palillos para higiene dental.
- Del estudio del cálculo dental se deduce el consumo de plantas y el procesamiento de materias primas vegetales. Los restos de polen y fragmentos de insecto son consistentes con un entorno boscoso. No hay evidencias de cocinado de alimentos (Hardy et al, 2016).
- El microdesgaste dental no oclusal presenta una alta densidad revelando el consumo de alimentos duros con desgaste por hueso triturado y arenilla, con un oportunismo alimenticio de amplio espectro que incluye carne y vegetales. Este patrón está presente también en los ejemplares de Homo antecessor (Pérez-Pérez et al, 2017).
- Un premolar inferior de leche.
- ATE 9-2, Una falange proximal del quinto dedo de la mano izquierda de un adulto. Robusta. (Carlos Lorenzo et al, 2014).
 |
| ATE 9-1 |
 |
| Hallazgo de la Sima del Elefante datado en 1,4 Ma, el resto más antiguo de Europa relacionado con homínidos |
Otros yacimientos
La mayor parte, corresponden a un periodo de mejora climática marcada por el subcrón Jaramillo (1,07-0,99 Ma).
- En Lezignan-la-Cébe (Hérault), Francia hallazgos líticos datados en 1,57 Ma.
- En el nivel IV del Alto de las Picarazas (Andilla y Chelva, Valencia, España) con una datación de 1,5 Ma han aparecido herramientas líticas del Modo I en sílex y cuarcita, y además restos de fauna (caballos y bisontes) con marcas de corte. Huesos de conejo muestran, además de las marcas de corte con útiles líticos, marcas de dentición humana. De confirmarse la existencia de huesos quemados, estaríamos ante la evidencia de la producción del fuego más antigua de la Península Ibérica.
- Kurgan Cimbal, Bogatiry y Rodniki, en Rusia. 1,2-0,8 Ma. Shchelinsky et al, 2003.
- En el Valle Medio del Loira, Francia (Despriée et al, 2010):
- Pont-de-la-Hulauderie, Saint-Hilaire-la-Gravelle. Hallazgos líticos de difícil datación, pero que podrían tener más de 1 Ma.
- En Pont-de-Lavaud, Francia, hallazgos líticos datados en 1,1 Ma.
- La Chaudronnière
- En L’Usine de Rosières à Lunery, Francia, hallazgos líticos datados en 1,1-1 Ma.
- En Untermassfeld, Alemania, hallazgos líticos datados en 1,07 Ma (Landeck y García, 2016).
- El yacimiento lítico de Soleihac (Haute-Loire, Francia) está datado en 1,07-0,99 Ma.
- En Grotte du Vallonnet, (Roquebrune-Cap Martin, Alpes Marítimos, Francia) hallazgos líticos datados en 1,05-1 Ma (de Lumley et al, 1988).
- En los niveles TD3-TD4 de La Gran Dolina (Atapuerca) aparecieron algunas herramientas líticas (1 Ma). En 2013, se encontró un "chopper" de calcita tallado en uno de sus lados con el fin de cortar o descarnar animales.
- En el yacimiento del Barranco de la Boella (La Canonja, Tarragona), datados en 1 Ma, han aparecido diferentes restos líticos y huesos de animales.Entre los restos líticos aparecen dos hachas trabajadas por las dos caras, lo que significaría que estamos ante el achelense más antiguo de Europa.
- En El Pino, Carrascosa del Campo, Cuenca, se ha comunicado el hallazgo de núcleos de cuarcita de cerca de 1 Ma.
- Happisburgh, Norfolk, Inglaterra, es el primer asentamiento conocido al norte del paralelo 45. Hallazgos líticos y huellas humanas (Nick Ashton et al, 2014). Datado en 0,99-0,78 Ma, cuando la isla estaba unida al continente. Descubierto por Simon Parfitt et al (2010). Para Chris Stringer, los autores quizá pertenecieran al Homo antecessor.
- Dursunlu, Turquía. 0,99-0,78 Ma. Gülec et al, 2009.
- Korolevo, Ucrania. 0,99-0,96 ka. Koulakovska et al, 2010.
- En Vallparadís (Terrasa, Barcelona) se han encontrado herramientas modo 1 bipolares olduvaienses datadas en 0,9 Ma, con restos de fauna asociados. Algunos restos de herbívoros presentaban marcas de corte y fracturaciones antrópicas, demostrando así un acceso primario de los humanos a las carcasas de los animales en relación al resto de grandes carnívoros.
 |
| Primeros poblamientos humanos de Iberia. Pueden identificarse dos dispersiones faunísticas diferentes. Gibert et al, 2015. |
- En Cueva Negra, Caravaca de la Cruz, Murcia, apareció tecnología achelense datada en 0,9 Ma, con restos de fauna quemada y trozo de sílex deformado por el fuego. Son los restos de fuego controlado por humanos más antiguos de Europa.
- Cueva Victoria, Cerro de San Ginés de la Jara, a 3 km del Mar Menor y a 13 km al este de Cartagena. Hallazgos datados entre 0,99-0,78 Ma.
- CV-0, falange.
- CV-1 y CV-2, fragmentos de húmero.
- En Ca'Belvedere di Monte Poggiolo (Forli, Emilia Romagna, Italia), hallazgos líticos datados en 0,85 Ma. Arzarello y Peretto, 2010.
- Dorn-Dürkheim, Alemania, DD31 más de 0,8 Ma. Hallazgos líticos.
- Pakefield, Inglaterra, artefactos de sílex datados en 0,7 Ma.
- En Cúllar-Baza 1 (Granada, España), una escasa colección de piezas de sílex, y en menor medida de cuarcita, que se enmarcan dentro de la tecnología Modo I-Olduvayense datada entre 0,78-0,65 Ma.
- En Solana del Zamborino, Granada, España, industria lítica en su mayor parte realizada con cuarzo y cuarcita aunque también se encuentran instrumentos de sílex, entre los que destaca un gran bifaz desconocido hasta el momento en Andalucía. En 2009 Gary Scott y Luis Gilbert realizaron un estudio geocronológico en el Berkeley Geochronology Center, publicado en la revista Nature, con el que descubrieron que el yacimiento era más antiguo de lo que se creía, situando su cronología de 0,76 Ma.
Sin datación precisa, en la Península Ibérica, disponemos además de los siguientes yacimientos:
- El Aculadero (Puerto de Santa María, Cádiz).
- El Rompido, Huelva.
- Magoito, Mironço, Aldeia Nova y Açafora, en los alrededores de Lisboa.
- Peteira (Tuy), Tomiño-Campo de Fútbol (Pontevedra), Chan de Vide – “Minas del Condado 3 y 5” (Pontevedra), Montes de Oleiros (Salvaterra de Miño y de As Neves), en las terrazas del tramo final del río Míño.
- Monfarracinos y Toro, en las terrazas del Duero.
- Hallazgos aislados en el entorno de Valladolid, en las terrazas del Pisueerga.
- Perilla del Castro y Betrocino, en las terrazas del Esla.
- Hallazgos en las terrazas del Tormes.
- Pueblanueva, en las terrazas del Tajo.
- Mas d’en Galí (Medinyà, Girona), Costa Roja (Sant Julià de Ramis, Girona) y el conjunto de Puig d’en Roca (Girona), en el curso medio del Ter.
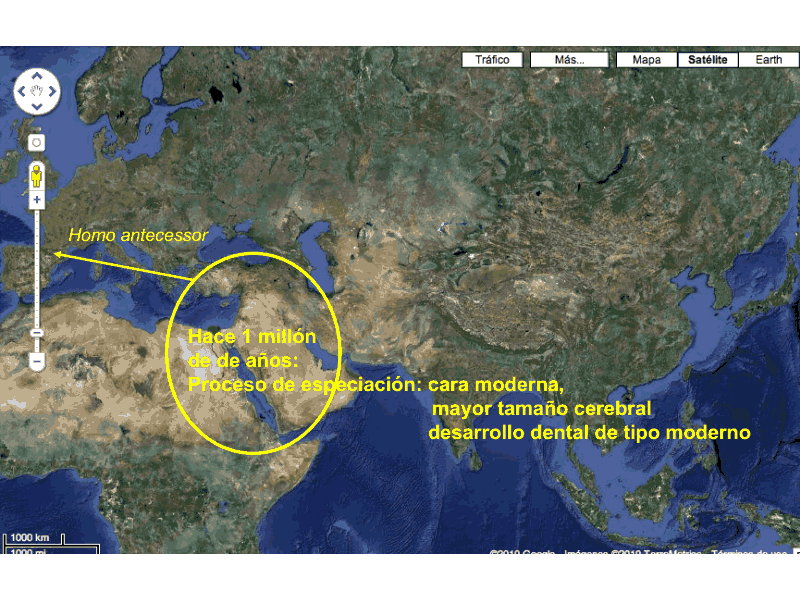 |
| Especiación de Homo antecessor. José María Bermúdez de Castro, 2013. |
Homo Antecessor, Bermúdez de Castro et al, 1997
Yacimiento de La Gran Dolina
A partir de la realización de una cata de 7 m2 en 1976, se recogen de la Gran Dolina de Atapuerca (Burgos) restos que alcanzan más de 0,9 Ma (J.M. Parés et al, 2013). El nivel 6, del que proceden los hallazgos, alcanza los 2,5 metros de espesor y está formado por una colada con escasa matriz arcillosa. TD6-2, el estrato Aurora (nombre de la descubridora Aurora Martín Nájera), contenía ca 170 fósiles de Homo antecessor correspondientes a 10-15 individuos casi todos menores de quince años; la mayoría menores de dos años con algunos bebés.
Dataciones por ESR proporcionaron una edad entre hace 0,77-0,85 Ma para el límite superior de TD6 y de 0.91 ± 0.25 Ma para el límnite inferior de TD4. Análisis de lumniscencia, proporcionaron una edad de TD6 de hace 846 ± 57 ka. La combinación de datos obtenidos por ESR, luminiscencia, bioestratigrafía y resultados paleomagnéticos soportan una edad post-Jaramillo para la capa TD4 de Gran Dolina (Claudia Álvarez-Posada et al, 2018). La datación directa por ESR del diente ATD6-92 ha producido unas fechas entre hace 0,949-0,772 ka (Duval et al, 2018).
Sobresalen:
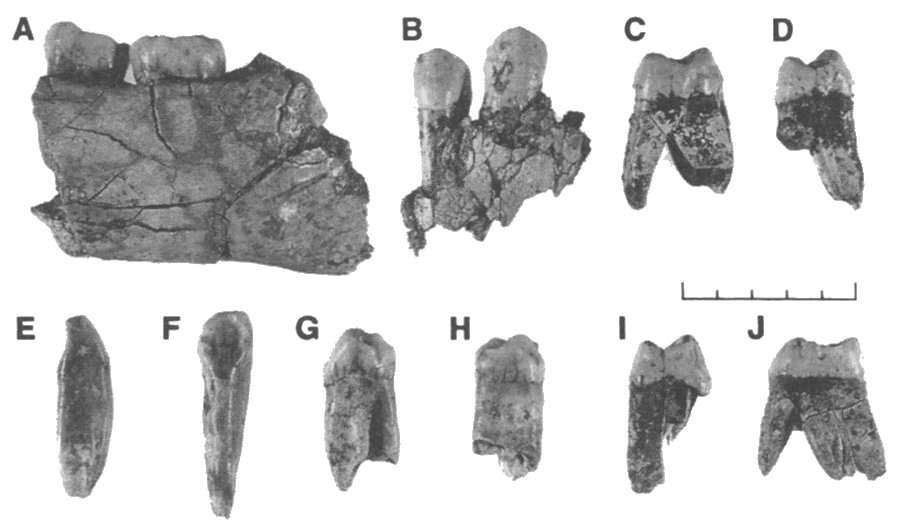 |
| ATD6-5 y dientes asociados. Carbonell et al, 1995. |
 |
| ATD6-96. Vista oclusal y lateral. |
- ATD6-5. Holotipo. Media mandíbula con sus tres muelas de un individuo joven. M3 está saliendo torcida, lo que debió originar dolor de muelas e incapacidad de morder. Sin los rasgos característicos de los preneandertales y pocos rasgos parecidos a los del ergaster.
- ATD6-15. Restos parciales de la cara de un niño entre 10 y 11,5 años de edad. (Bermúdez de Castro et al 1997). Conserva el torus derecho y parte de la región de la glabela. Senos frontales muy desarrollados. La capacidad craneal pudo llegar a 1.000 cc.
- ATD6-19. Pequeño fragmento cigomaxilar de un adulto, con un tubérculo cigomaxilar de las mismas características que el de ATD6-58, que se proyecta hacia fuera 2 mm (Arsuaga et al, 1999).
- ATD6-22. Rótula derecha.
- ATD6-38. Muy similar en tamaño y forma a ATD6-69. Su topografía sugiere claramente la presencia de una fosa canina (Arsuaga et al, 1999).
- ATD6-43, fragmento de radio casi completo. El índice braquial y las proporciones son más próximos a los del ergaster y a los de los humanos modernos que a los de preneandertales y neandertales.
- El ATD6-50 clavícula muy larga y relativamente delgada, con una curvatura del eje pronunciada y epífisis relativamente pequeñas, similar a la de los neandertales (Carretero et al, 1999).
- ATD6-56. Rótula izquierda asociada a la rótula derecha ATD6-22. Presenta evidencia de un traumatismo.
- ATD6-58. Fragmento cigomaxilar izquierdo de un adulto. Exhibe una notable fosa canina así como un gran tubérculo cigomaxilar situado en posición maxilar que se proyecta hacia fuera 3,3 mm (Arsuaga et al, 1999).
- ATD6-69, El Chico de la Gran Dolina (indviduo 3). Gran parte del lado izquierdo de la cara, excepto las apófisis frontales de los huesos cigomáticos y maxilares. Conserva I2 derecho y P3, M1, M2 y M3 izquierdos permanentes. C y P4 derechos estaban finalizando el proceso de erupción. M2 izquierdo, todavía incluido en su cripta mientras que el germen de M3 se mantuvo unido al hueso por debajo de la cripta de M2. La edad dental, de acuerdo con los estándares modernos, se ha estimado en 10-11 años. Morfología totalmente moderna, incluyendo el patrón de desarrollo dental (Bermúdez de Castro et al, 1999), con rasgos más cercanos a los de los cráneos de Zhoukoudián y Dali y a los de Cromañón que a los neandertales (Rodrigo S. Lacruz et al, 2013, Sarah E. Freidline et al, 2013).
- ATD6-92, fragmento de diente de un varón. Sobre este ejemplar se efectuó la primera datación por ESR de antecessor (Duval et al, 2018) y se obtuvo el proteoma humano más antiguo hasta la fecha (Welker et al, 2020).
- ATD6-93, M1 inferior derecho que exhibe el mayor diámetro bucolingual registrado hasta ahora en Homo (José María Bermúdez de Castro et al, 2017).
- ATD6-95 y ATD6-70+107 astrágalo izquierdo casi completo y segundo metatarsiano derecho asignados tentativamente al mismo homínido, un macho adulto. El astrágalo es largo y alto, con una tróclea larga y ancha y con un cuello proporcionalmente corto.
- ATD6-96. Fragmento izquierdo de la mandíbula de una mujer muy joven. Exhibe un tubérculo pterigoideo hipertrofiado, una característica incluida en la lista de apomorfías neandertales (Carbonell et al, 2005; Rak et al, 1994; Weaber, 2009).
- ATD6-121. Húmero infantil.
- ATD6-124. 3/4 partes de un cuarto metatarsiano derecho. Según L. Martín-Francés et al (2013) el hueso estaba roto y se estaba soldando en el momento del fallecimiento del individuo. La presencia de esta fractura sugiere que la correspondiente población recorría grandes distancias o se desplazaba frecuentemente por un terreno desigual.
- ATD6-148. Rafa. Húmero izquierdo de adulto muy joven vigoroso. Quebrado para obtener la médula. Marcas de descarnado y destrozo de los cóndilos para separar las masas musculares. Al igual que ATD6-121, exhibe una gran fosa del olécranon y unos pilares laterales y mediales muy finos, similares a los de los homínidos del Pleistoceno Medio europeo, los neandertales y Bodo (Carretero et al, 2009; Bermúdez de Castro et al, 2012).
- ATD6-100/168. Un gran fragmento de parietal, el mayor recuperado hasta el momento del neurocráneo de antecessor. La morfología de las fracturas es compatible con un trauma peri-mortem. Las zonas parietales superiores son planas. Están presentes un gran foramen parietal y un foramen accesorio pequeño. Los vasos meníngeos medios no están particularmente desarrollados, pero se distribuyen en ambos sectores anterior y posterior, con una vascularización parietal procedente de la rama posterior. Los vasos meníngeos muestran múltiples conexiones de menor importancia con los sistemas pericraneal y diploico. El diploe no está particularmente desarrollado, y no se detectan grandes canales diploicos. El hueso es delgado. Esta descripión sugiere que el hueso perteneció a un juvenil. Bruner et al, 2016.
También, más de 800 artefactos líticos:
- En el nivel TD4, un chopper que fue utilizado para descuartizar huesos de animales, datado en 1 Ma. El techo del nivel TD4 había deparado ya varias herramientas de cuarcita manufacturadas a partir de cantos del río Arlanzón.
- En TD5, tres artefactos, entre ellos un núcleo de cuarcita y una lasca de sílex.
- En TD6, asociada a los restos humanos, aparece una abundante tecnología lítica (n>300) con núcleos y lascas (industrias arcaicas o Modo I); la materia prima utilizada es el sílex, cuarzo, cuarcita, caliza y arenisca.
- Pese a que la tecnología no varió sustancialmente existieron dos fases principales de ocupación humana: En una primera, los Homo utilizaron muy marginalmente la cavidad pero poco a poco su presencia se va haciendo más intensa hasta que en las fases medias y finales se asienta un campamento base, en el que se incrementan notablemente las actividades, incluyendo los eventos de canibalismo (Marina Mosquera et al, 2018).
En cuanto al esqueleto postcraneal, H. antecessor se parece más a los HAM que a los homínidos europeos de mediados y finales del Pleistoceno (Carretero et al, 1999; Lorenzo et al, 1999), aunque algunos rasgos también se comparten con estos últimos (Pablos et al, 2012). Los metatarsianos no difieren significativamente de los correspondientes a los humanos modernos, neandertales y ejemplares de la Sima de los Huesos.
En resumen, H. antecessor se caracteriza por una combinación única de características plesiomórficas y derivadas (Bermúdez de Castro et al, 2017).
- Plesiomórficas:
- La dentición: desarrollo de un cíngulo en premolares y molares, crenulaciones del esmalte profusas, asimetría de la corona del primer premolar inferior, gran tamaño en molares y sistemas radiculares complejos y robustos. Ausencia de espacio retromolar. M3 parcialmente cubierto por la rama mandibular. Fuerte tubérculo molar en el dm1 deciduo. Presencia de cúspides mesial y lingual en el canino superior deciduo. Las caras de los premolares mandibulares muestran crestas y surcos marginales mesiales y distales, que se conectan con el cíngulo. La forma de la corona del P3 mandibular es fuertemente asimétrica, con un notable talónido y un gran polígono oclusal. El P4 mandibular muestra un contorno asimétrico, con un metacónido desplazado mesialmente con talónido abultado y un amplio polígono oclusal. P3 y P4 mandibulares muestran un sistema radicular complejo, con una raíz mesiobucal con dos canales de pulpa y una raíz distolingual con un solo canal. M1<M2 para las series molares superior e inferior. M1 y M2 mandibulares exhiben un patrón Y de los surcos bucales y linguales que separan las cinco cúspides principales. El esmalte de la superficie oclusal de los dientes poscaninos está moderada o notablemente crenulado. Cuello radial largo tanto desde un punto de vista absoluto como relativo.
- Los rasgos endocraneales de los parietales.
- Frente huidiza y toro supraorbital de doble arco.
- El proceso estiloides esta fusionado al basicráneo.
- Suelo nasal inclinado.
- Ausencia de mentón.
- Posición del foramen mental al nivel de P3-P4 mandibular.
- Baja posición de la línea milohioidea en relación al margen alveolar en el nivel M3 mandibular. Trayectoria paralela de la línea milohioidea en relación al margen alveolar.
- Fosa pterigoidea poco profunda.
- Apomorfias compartidas con Hominini posteriores.
- Torus supraorbital doblemente arqueado.
- Los parietales muestran un aplanamiento parasagital de las superficies superiores.
- Posición de la prominencia lateral al nivel del talónido del M2 mandibular. Ausencia de prominencia alveolar. Trigonum postmolare inclinado oblicuamente. Fosa masetérica poco profunda. Fosa subalveolar moderadamente profunda. Perfil regular del gonion. El ratio longitud/anchura de la arcada alveolar mandibular está claramente por debajo de cien.
- Incisivos permanentes mandibulares bucolingualmente expandidos con respecto a H. habilis s.s. y otros Hominini africanos y euroasiáticos del Pleistoceno Temprano. P3>P4 para el área coronal de los premolares. Dientes poscaninos más pequeños que los de H. habilis s.s. y en el rango de H. ergaster, H. erectus s.l., y Hominini europeos del Pleistoceno Medio. M3 mandibular notablemente reducido con respecto a M1 y M2.
- Tuberosidades radiales enfrentadas anteromedialmente.
- El patrón de remodelación facial en la ontogenia.
- La ontogenia dental.
- Características derivadas, compartidas con los neandertales y los HAM.
- Borde superior convexo de la escama del temporal, y una posición anterior del conducto incisivo, que es casi vertical (Arsuaga et al, 1999).
- Planum alveolare ausente o débil.
- Capacidad craneal, estimada en más de 1.000 cc.
- Características derivadas, compartidas con los HAM.
- La cara. Más moderna que la de los homínidos africanos del Pleistoceno Inferior, caracterizados por un prognatismo pronunciado. Es plana, con un nasal saliente, maxilar poco proyectado y mandíbula delgada. Fosa canina, consecuencia de una reorientación hacia delante y abajo de las placas infraorbitarias cuyo borde inferior parte de una posición horizontal y alta.
- Posición medial del punto más profundo de la hendidura mandibular.
- Rótula relativamente estreha, con indices rotulianos altos.
- Características derivadas compartidas con los neandertales y la población de la Sima de los Huesos.
- Presencia de un tubérculo pterigoideo medial.
- Polígono oclusal reforzado y comprimido y un contorno externo sesgado de la M1 permanente con protuberancia abultada del hipocono.
- La región mastoidea es una reminiscencia de la de los neandertales.
- Incisivos superiores permanentes muestran forma de pala, especialmente I2.
- Presencia de cresta media del trigónido.
- Fosa olécranona grande y pilares medianos y laterales muy finos
- Clavícula muy larga en términos absolutos, relativamente delgada (bajo índice de robustez), con pronunciada curvatura del eje y epífisis relativamente pequeña.
- Tróclea del astrágalo relativamente estrecha.
MORFOLOGÍA FACIAL
|
ESCAMA TEMPORAL
| |
Homo habilis
Homo rudolfensis
|
Cara plana o cóncava.
Placas infraorbitales dispuestas coronalmente, verticales u orientadas hacia arriba.
La apertura nasal no se sitúa por delante de las placas infraorbitarias.
|
Borde bajo y rectilíneo.
|
Grado erectus
|
Cavidad nasal ligeramente más adelantada respecto a las placas infraorbitarias. Prognato.
Huesos nasales dispuestos más horizontalmente.
Zona de crecimiento óseo bajo la nariz. Osteoblastos. Topografía subnasal plana. |
Borde bajo y rectilíneo.
|
Homo antecessor
|
Placas infraorbitales orientadas coronalmente e inclinadas hacia abajo, con fosa canina.
Resorción ósea bajo la nariz. Osteoclastos. Topografía subnasal convexa. Tubérculo pterigoideo medial hipertrofiado. |
Borde elevado y arqueado.
|
Homo neanderthalensis
|
Cara de aspecto apuntado.
Cavidad nasal situada más hacia delante que en las demás especies.
Huesos nasales dispuestos horizontalmente.
A ambos lados de la nariz, las paredes de la cavidad nasal y las placas infraorbitarias forman una única superificie sin fosa canina, orientadas oblicuamente.
Tubérculo pterigoideo medial hipertrofiado. |
Borde elevado y arqueado.
|
Homo sapiens
|
Con fosa canina.
Resorción ósea bajo la nariz. Osteoclastos. |
Borde elevado y arqueado.
|
 |
| Homo antecessor. ATD6-69 y ATD6-15. |
Estatura, proporciones y peso
Una gran mayoría de individuos medirían entre 160 y 180 cm, siendo el promedio de los machos algo superior al promedio de las hembras, como sucede en la actualidad en nuestra especie. En función de la tibia, el fósil original que se exhibe en el MEH, mediría unos 175 cm. Proporciones corporales: similares a las de Homo sapiens, pero con el tronco más ancho. Peso corporal: un 30% mayor que sapiens, debido a una mayor masa esquelética y muscular y a una mayor anchura del tronco.
Las diferencias de peso y estatura entre machos y hembras serían similares a las del sapiens.
Longevidad: menor que la de Homo sapiens. Su estilo de vida no permitiría una vida más allá de los 40 o 45 años. La esperanza de vida al nacimiento sería de entre quince y veinte años.
Posibles causas de muerte: parto, infecciones de heridas y de la cavidad bucal, traumatismos producidos por golpes accidentales o por violencia intra-específica, predadores.
La industria lítica hallada es pre-achelense, Modo 1 o olduvaiense evolucionado, con ausencia de hachas de mano. El conjunto lítico es rico y diversificado. Los bloques de sílex para núcleos fueron amontonados de forma intencional. Se observa la representación completa de la cadena operativa que exige la fabricación de los instrumentos líticos, es decir, la existencia de núcleos, lascas, útiles, así como de fragmentos producto de la actividad de talla. La actividad productiva de taller en el sitio se ha confirmado mediante diversos remontajes. Predominan las bases positivas, de las que son minoría las retocadas. Se ha comprobado su utilización para cortar carne, preparar huesos para romperlos, descortezar y raer madera y posiblemente se utilizasen también para tratar la piel. Se han documentado actividades de caza y los primeros rastros de canibalismo en la prehistoria (Andréu Ollé et al, 2013)
Las diferencias de peso y estatura entre machos y hembras serían similares a las del sapiens.
Ciclo de vida
Parto similar al de Homo sapiens en cuanto a la fisiología, movimientos del niño y modo de presentación, pero con un tránsito más holgado por el canal del parto, debido a las mayores dimensiones de la pelvis. Lactancia: entre tres y cuatro años. Desarrollo probablemente casi tan largo y complejo como en nuestra especie, con infancia, niñez, fase juvenil y adolescencia, incluido un estirón puberal de intensidad similar a la nuestra.Longevidad: menor que la de Homo sapiens. Su estilo de vida no permitiría una vida más allá de los 40 o 45 años. La esperanza de vida al nacimiento sería de entre quince y veinte años.
Posibles causas de muerte: parto, infecciones de heridas y de la cavidad bucal, traumatismos producidos por golpes accidentales o por violencia intra-específica, predadores.
Modo de vida y dieta
Se trata de una asentamiento de larga duración (un campamento base), pero la antropofagia sugiere una cierta sincronicidad en la deposición, de unas decenas o cientos de años. Campaña et al (2016) proponen un origen alóctono: la actividad de los Homo se habría desarrollado fuera de la cueva, pero muy cerca de ella. la excelente conservación tafonómica de los huesos indica un tiempo muy corto a la exposición externa, un transporte rápido y en distancias muy cortas. Una consecuencia de esta hipótesis es que la datación de los correspondientes fósiles podría ser más antigua de la que se supone actualmente.
Cazadores y recolectores de pequeños animales, huevos y todo tipo de plantas silvestres comestibles. Estos seres utilizaban distintas estrategias para cazar, se organizaban en clanes de entre 15 y 40 individuos jerarquizados y transportaban las piezas de diferentes formas, seleccionándolas o no según su tamaño, la hora del día, la distancia a la base o la presencia de otros carnívoros en el entorno. De estos hechos podemos inferir una gran complejidad social con reparto de tareas (Palmira Saladié, 2011). Contactos esporádicos con otros grupos para evitar la endogamia. La presencia de costillas (que son consumidas por los grandes depredadores) y de marcas de corte realizadas con cuchillos de piedra permite deducir que mataban ellos mismos a estos animales. Los restos humanos presentan las mismas huellas de tratamiento con herramientas de piedra que los restos de animales de lo que se deduce canibalismo (Palmira Saladié et al, 2015). Según Palmira Saladié et al (2012) los homínidos de TD6 realizaban ataques de bajo riesgo a los miembros de otros grupos para defender el acceso a los recursos dentro de sus propios territorios y tratar de ampliar sus territorios a costa de los grupos vecinos. Según Guillermo Rodríguez-Gómez et al (2013), el entorno era muy rico en recursos, con un nicho vacío para los predadores, y los homínidos podían aprovisionarse de forma continua de abundante carne y grasa por lo que hay que descartar el hambre como causa del canibalismo. Podría tratarse de víctimas de enfrentamientos territoriales de carácter estructural.
Rosa Huguet el al (2013) han estudiado los restos de fauna recuperados en La Gran Dolina (niveles 3-4 y 6) y en la Sima del Elefante (niveles 9-14). Abundan los restos de fauna como producto de la práctica cinegética, con abundantes marcas de corte en los restos esqueléticos, que han sido fragmentados sistemáticamente por la acción antrópica. Las estrategias de subsistencia tuvieron un éxito suficiente como para permitir que los grupos de homínidos sobreviviesen y se reprodujesen en número suficiente. Por lo tanto, estos primeros seres humanos tendrían la capacidad de mantener una ocupación continua de Europa.
Otras conclusiones:
Campamentos con cabañas de madera y quizás pieles de animales, pero también uso de abrigos y vestíbulos de cuevas.Cazadores y recolectores de pequeños animales, huevos y todo tipo de plantas silvestres comestibles. Estos seres utilizaban distintas estrategias para cazar, se organizaban en clanes de entre 15 y 40 individuos jerarquizados y transportaban las piezas de diferentes formas, seleccionándolas o no según su tamaño, la hora del día, la distancia a la base o la presencia de otros carnívoros en el entorno. De estos hechos podemos inferir una gran complejidad social con reparto de tareas (Palmira Saladié, 2011). Contactos esporádicos con otros grupos para evitar la endogamia. La presencia de costillas (que son consumidas por los grandes depredadores) y de marcas de corte realizadas con cuchillos de piedra permite deducir que mataban ellos mismos a estos animales. Los restos humanos presentan las mismas huellas de tratamiento con herramientas de piedra que los restos de animales de lo que se deduce canibalismo (Palmira Saladié et al, 2015). Según Palmira Saladié et al (2012) los homínidos de TD6 realizaban ataques de bajo riesgo a los miembros de otros grupos para defender el acceso a los recursos dentro de sus propios territorios y tratar de ampliar sus territorios a costa de los grupos vecinos. Según Guillermo Rodríguez-Gómez et al (2013), el entorno era muy rico en recursos, con un nicho vacío para los predadores, y los homínidos podían aprovisionarse de forma continua de abundante carne y grasa por lo que hay que descartar el hambre como causa del canibalismo. Podría tratarse de víctimas de enfrentamientos territoriales de carácter estructural.
Rosa Huguet el al (2013) han estudiado los restos de fauna recuperados en La Gran Dolina (niveles 3-4 y 6) y en la Sima del Elefante (niveles 9-14). Abundan los restos de fauna como producto de la práctica cinegética, con abundantes marcas de corte en los restos esqueléticos, que han sido fragmentados sistemáticamente por la acción antrópica. Las estrategias de subsistencia tuvieron un éxito suficiente como para permitir que los grupos de homínidos sobreviviesen y se reprodujesen en número suficiente. Por lo tanto, estos primeros seres humanos tendrían la capacidad de mantener una ocupación continua de Europa.
Otras conclusiones:
- Se constata un acceso primario de los homínidos (anterior al de los carroñeros) y no se ha constatado acceso secundario (Palmira Saladié et al, 2014)
- Se efectuaban todas las tareas de carnicero.
- No se seleccionaban los animales.
- Los homínidos desarrollaron un comportamiento complejo de estrategias de subsistencia oportunistas y generalistas con un vasto conocimiento y aprovechamiento del territorio.
La industria lítica hallada es pre-achelense, Modo 1 o olduvaiense evolucionado, con ausencia de hachas de mano. El conjunto lítico es rico y diversificado. Los bloques de sílex para núcleos fueron amontonados de forma intencional. Se observa la representación completa de la cadena operativa que exige la fabricación de los instrumentos líticos, es decir, la existencia de núcleos, lascas, útiles, así como de fragmentos producto de la actividad de talla. La actividad productiva de taller en el sitio se ha confirmado mediante diversos remontajes. Predominan las bases positivas, de las que son minoría las retocadas. Se ha comprobado su utilización para cortar carne, preparar huesos para romperlos, descortezar y raer madera y posiblemente se utilizasen también para tratar la piel. Se han documentado actividades de caza y los primeros rastros de canibalismo en la prehistoria (Andréu Ollé et al, 2013)
Del conjunto faunístico asociado se deduce un ambiente de bosque abierto o con pradera en proximidad y un remanso o laguna muy cercano, con un clima cálido, de humedad estacional; unas condiciones climáticas similares a las de los actuales inviernos en la zona. En este tiempo, la cavidad de la Gran Dolina fue un lugar de consumo.
Los restos de polen sugieren una recuperación de
temperaturas después de una fase fría.
La clasificación de los restos de La Gran Dolina.
Lo habitual para los restos europeos de transición era
clasificarlos como heidelbergensis.
Para Carbonell (1995), se trataría de un taxón diferente al erectus (que se extingue) y al ergaster (que daría lugar al sapiens).
Howell le asignó el paleodemo Atapuerca-Gran Dolina.
El equipo de Atapuerca propuso la nueva especie Homo antecessor (Carbonell et al, 1997), con una topografía mediofacial completamente moderna y otros
rasgos que lo apartaban de erectus,
ergaster y neanderthalensis.
- Está relacionada con los pobladores del Pleistoceno antiguo de China. Ambos demos procederían del grupo que abandonó África hace 1,9 Ma. Para comprobar la relación sería necesario hallar fósiles de este periodo en el este de Europa y oeste de Asia.
- En contra de la asignación a una nueva especie, hay que mencionar lo fragmentario de los restos y que corresponden en su mayoría a individuos infantiles (David Rabadà, artículo en Catalunya Vanguardista, 2013).
María Martinón-Torres et al (2007) a partir de un estudio fenétido y cladístico de la dentición diferencian entre:
- Un bloque africano (A. africanus, A. afarensis, H. habilis, H. ergaster y H. mauritanicus).
- En la dentición posterior, aumento significativo de cúspides accesorias y rasgos que complican profusamente la superficie oclusal.
- Un bloque euroasiático (H. antecessor, H. erectus s.s., H. heidelbergensis y H. neanderthalensis).
- En la dentición anterior, forma en pala triangular, con una convexidad labial pronunciada y una superficie oclusal en "V" por el desarrollo de un surco estrecho secundario.
- En la posterior, rasgos derivados de la reducción dental como la pérdida de cúspides en molares, simplificación de las superficies oclusales y aparición de patrones de surco diferentes del driopitecino. Cresta media del trigónido continua con una fovea anterior corta y profunda. Cresta transversa continua en segundos premolares inferiores.
- H. sapiens y H. georgicus muestran una posición intermedia.
Para Emiliano Aguirre (2008) no
tendría relación filogenética directa ni con erectus ni con heidelbergensis
o sapiens. Aguirre (2000) es defensor de la teoría mutirregional de
evolución hacia el sapiens y cree que
rasgos faciales compartidos por antecessor,
erectus y sapiens indican un
aporte genético muy variado como
origen de los humanos actuales.
El rostro de antecessor combina características compartidas con el HAM con otras similares a las de los neandertales y grado heidelbergensis. Sarah E. Freidline et al (2013) han sugerido que algunos de los rasgos faciales que caracterizan a los últimos humanos modernos pueden haberse desarrollado varias veces en la evolución humana.
El equipo de Atapuerca considera a Homo atecessor un buen candidato a la ascendencia común de HAM y neandertales. Mendez et al (2016) han estimado que la separación entre neandertales y HAM se produjo hace 806-447 ka para el ADN-Y. Esta fecha cae dentro del rango de antecessor y fuera de heidelbergensis. Gómez-Robles et al (2013) señalan que ninguna especie conocida muestra una morfología dental compatible con la que se podría esperar para un antecesor común de neandertales y HAM.
José María Bermúdez de Castro y María Martinón-Torres examinan la hipótesis de la convergencia de Sarah E. Freidline et al (2013) a la luz de los fósiles humanos recuperados en TD6. Destacan que basan su interpretación en la ausencia de una cara totalmente moderna en el Pleistoceno Medio africano, pero debido a la ausencia de fósiles, solo es posible tener en cuenta Kabwe y Bodo. Para este último, algunas características del infraorbitario permitirían alinearlo con H. sapiens. La falta de una muestra representativa, llama a la prudencia.
Como una alternativa a las interpretaciones de Sarah E. Freidline et al (2013), los autores proponen una cladogénesis del género Homo durante el Pleistoceno Temprano, que dio lugar a un aumento del tamaño craneal y a una morfología facial moderna.
- KNM-WT 15000 representa la condición primitiva del clado Homo
- H. antecessor deriva probablemente de este cladogénesis y representa una rama lateral confinada a Europa Occidental (Bermúdez de Castro et al, 2003; Carbonell et al, 2005; Martinón-Torres et al, 2007; Bermúdez de Castro y otros, 2012). H. antecessor comparte características derivadas con neanderthalensis, la población de la Sima de los Huesos, los Homo del Pleistoceno Medio de China y sapiens y estaría muy cerca de su ancestro común más reciente. Precisamente debido a esta posición filogenética, niegan una posible relación ancestro-descendiente con la especie ñ (José María Bermúdez de Castro et al, 2013; José María Bermúdez de Castro et al, 2014; José María Bermúdez de Castro et al, 2015).
- El prognatismo mediofacial que puede observarse en los neandertales clásicos (Rak, 1986), y que también está presente en los homínidos del Pleistoceno de Europa Oriental (Arsuaga et al, 1999), representaría otra condición derivada que podría haber surgido a partir de una cara similar a la moderna (Arsuaga et al, 1999). Estos rasgos fueron retenidos por los neandertales (sinapomorfías) (Gómez-Robles et al, 2007; Bermúdez de Castro y Martinón-Torres 2012; Martinón-Torres et al, 2013).
- El espécimen de Bodo presenta algunas sinapomorfias con neandertales y HAM en diversos grados y podría formar parte de uno de los linajes de la cladogénesis que en África dio lugar a H. sapiens y en Europa a H. neanderthalensis.
 |
| Estratos y dataciones de La Gran Dolina (J.M. Parés et al, 2013). |
La cara del Homo antecessor
Más vídeos en Antena3
Otros yacimientos
Cueva Negra del Estrecho del Río Quípar (España) 0,78 Ma, según datos bioestratigráficos (M. J. Walker et al, 2012). Fragmentos de de húmero y cúbito humano y dientes.
En España hay yacimientos líticos datados en más de 0,75 Ma en Barranco León, Fuentenueva-3, Monfarracinos, Galisteo, Talavera, El Espinar, Atapuerca y Huéscar.
En España hay yacimientos líticos datados en más de 0,75 Ma en Barranco León, Fuentenueva-3, Monfarracinos, Galisteo, Talavera, El Espinar, Atapuerca y Huéscar.
Enlaces:
- Les vagues de peuplements humains au Pléistocène inférieur et moyen dans le bassin de la Loire moyenne, région Centre, France. Apports de l’étude des formations fluviatiles. Jackie Despriée et al (2009).
- Estratigrafía de la Gran Dolina.
- Origen del poblamiento en la Península Ibérica en el Pleistoceno Inferior: el registro fósil y lítico.
- David Sánchez
- El origen del poblamiento en la Península Ibérica en el Pleistoceno Inferior parte I: el registro fósil y lítico.
- El origen del poblamiento en la Península Ibérica en el Pleistoceno Inferior Parte 2: el registro europeo, asiático y las rutas de acceso.
- Entrevista a José María Bermúdez de Castro con ocasión del 18º aniversario de la publicación en Science del Homo antecessor.
- Roberto Sáez
Adrián Pablos: Deportistas de hace 500 ka.
Hola Juan Manuel, Theropithecus, del que ahora ya disponemos de 5 dientes (tenemos 4 nuevos que están en vías de publicación) solo se encuentra en Cueva Victoria. En Barranco León 5 así como en el resto de yacimientos de Orce (Fuentenueva 3, Venta MIcena, Barranco del Paso, etc.) no se encuentra este primate. Lo cual nos indica que debido a la diferencia de edades (Cueva Victoria 0,9 m.a. y los distintos yacimientos de Orce de 1,2-1,4 m.a.) pensamos que se trata de una dispersión diferente. La primera implicaría Homo, Hippopotamus antiquus y Equus altidens, y la segunda implicaría a Elephas antiquus y Theropithecus oswaldi. La presencia de Homo en Cueva Victoria no se sabe si responde a un desplazamiento desde Orce o bien a una nueva entrada desde África.
ResponderEliminarLa discusión sobre si realmente los restos de Atapuerca son una especie nueva o realmente son una subespecie de H. erectus todavía está abierta (así piensan paleoantropólogos como Tobias, Wang o Etler).
Francesc Ribot
Otra cosa, las últimas dataciones de Ceprano arrojan datos sorprendentes, y parece ser que no pasaría de 0,4 m.a., con lo cual habría que situarlo como un erectus evolucionado o, con mayor probabilidad, como un heidelbergensis.
ResponderEliminarFrancesc Ribot
Gracias, Fracesc Ribot. Importantes correcciones. En consecuencia con tu información, actualizo el post.
ResponderEliminarLa información sobre el yacimiento de Kozarnika indica una datación de 1.600.000-1.400.000 años. Luego indica industria achelense. ¿Tan temprano? Parece un error, los datos que circulan en internet indican niveles "no achelenses". No parece haber mucha más información sobre bifaces, parece un error. Saludos.
ResponderEliminarEfectivamente, se trata de un error que corrijo de inmediato. Muchas gracias.
EliminarEste comentario ha sido eliminado por el autor.
ResponderEliminar