 |
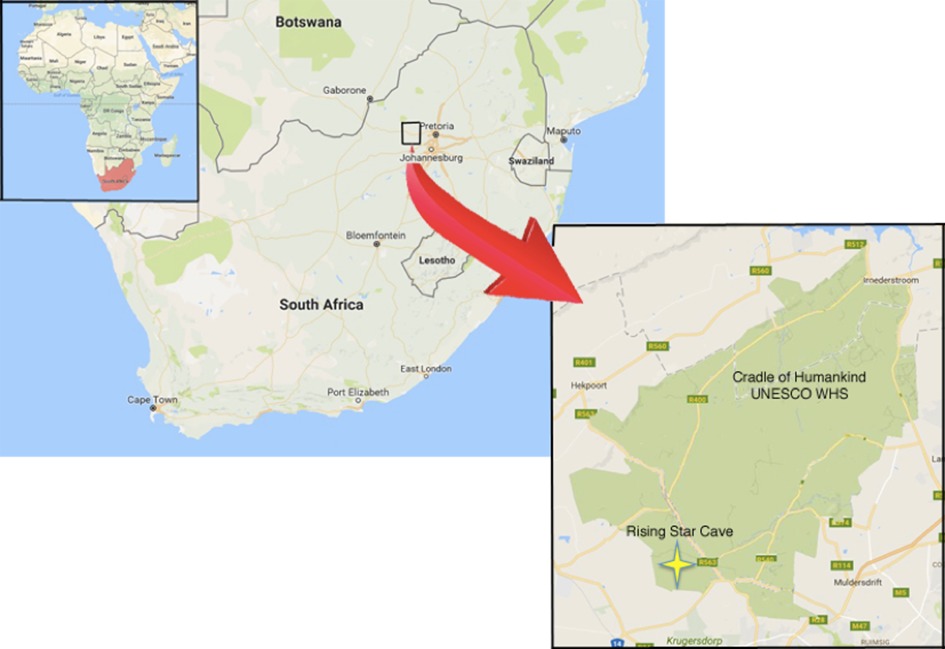
| Ubicación de Rising Star Cave. Hawks et al, 2017. |
 |
| Sistema de Cuevas Rising Star, con las localidades UW 101 (Dinaledi) y UW 102 (Lesedi). Hawks et al, 2017. |
 |
| Sistema de cuevas Rising Star. Kruger et al, 2016. |
 |
| Sección de Rising Star Cave |
Rising Star Cave es un sistema de cuevas localizado en el Cradle of Humankind World Heritage Site cerca de Krugersdorp en Sudáfrica, descubierto el 13 de septiembre de 2013 por los espeleólogos Steve Tucker y Rick Hunter, dentro de una exploración organizada por el geólogo Pedro Boshoff.
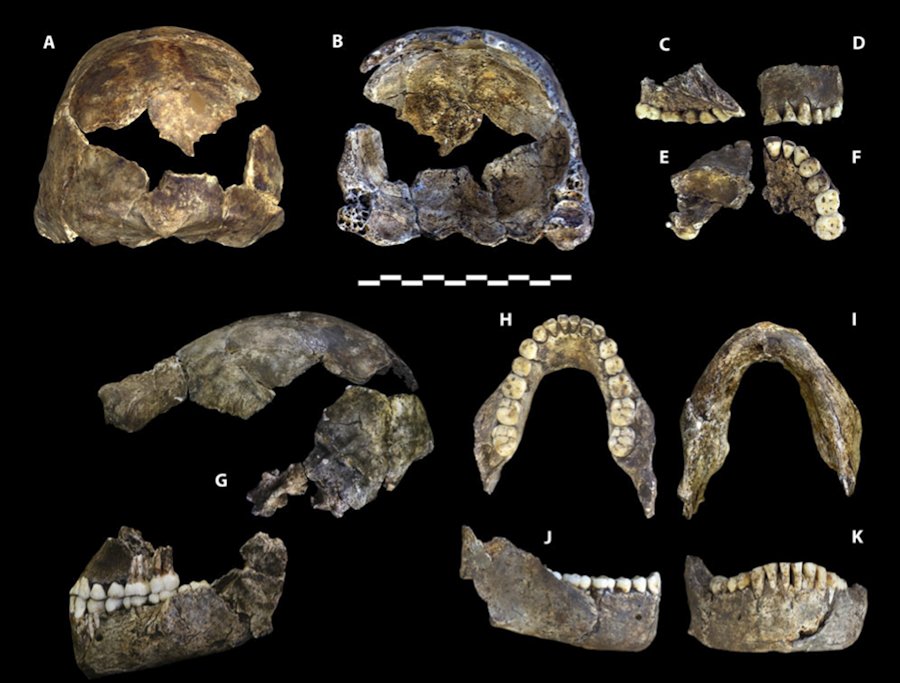 |
Holotipo Homo naledi. DH1. Formado por una calota parcial (U.W. 101-1473), un maxilar también parcial (U.W. 101-1277) y una mandíbula casi completa (U.W. 101-1261). (Berger, et al. 2015). |
El hallazgo fue notificado a Lee Berger el 1 de octubre de 2013 quien dirigió la excavación, financiada por National Geographic. La recuperación de los fósiles corrió a cargo de K. Lindsay Eaves, Marina Elliott, Elen Feuerriegel, Alia Gurtov, Hannah Morris, y Becca Peixotto, seleccionadas por su constitución ligera y experiencia paleoantropológica y espeleológica.
- Entre octubre de 2013 y marzo de 2014 se recuperaron en las unidades 2 y 3 de una cámara aislada (Cámara Dinaledi; Dirks et al, 2015) y se catalogaron 1.550 fósiles de un mínimo de quince individuos de todas las edades (8 inmaduros sobre 13 en los que ha podido determinarse la edad aproximada; Bolter et al, 2018) asignados a la nueva especie Homo naledi (Lee R. Berger et al, 2015). Naledi significa estrella en sesotho. Se trata del mayor conjunto de una especie de Hominini descubierto en África. Es probable que la cámara contenga más fósiles.
- 1.413 muestras de hueso, incluyendo huesecillos del oído interno y 137 muestras dentales aisladas; otros 53 dientes están presentes en muestras de hueso mandibular o maxilar.
- Posiblemente la cámara constituyese un lugar de enterramiento (pero Aurore Val, 2016; ver comentarios de Kate Wong, José María Bermúdez de Castro; réplica de Dirks et al, 2016, con comentarios de John Hawks; Egeland et al, 2018).
- Gran número de fósiles recuperados, que sugiere una deposición intencional.
- No hay rastro de depredadores ni de otros animales, salvo unos pocos huesos de aves y roedores, ni señales de catástrofe o flujo de agua. Los fósiles ocupan diferentes capas, lo que sugiere su deposición a lo largo de un rango de tiempo considerable.
- Los huesos no presentan marcas de corte.
- No hay evidencias de ocupación en toda la cueva Rising Star.
- Para acceder a la Cámara Dinaledi, debió utilizarse una entrada similar a la actual y traspasar estrechamientos inaccesibles a los mamíferos de tamaño medio. El trayecto debió requerir luz artificial. Los sedimentos de la Cámara Dinaledi son diferentes a los del resto de la Cueva, lo que indica que la cámara permaneció aislada a lo largo del tiempo.
 |
| Plano de la Cámara Lesedi. Hawks et al, 2017. |
En 2017 Hawks et al informaron de la recuperación de 133 fósiles adicionales en la Cámara Lesedi, otra cavidad dentro del sistema de cuevas Rising Star a 30 m por debajo de la superficie. No hay una ruta directa entre las cámaras Lesedi y Dinaledi. Como en el caso de los restos recuperados en Dinaledi, los fósiles estaban intactos y sin signos de haber quedado expuestos al ambiente de superficie. No hay evidencia de transporte, actividad de carnívoros, traumas peri-mortem o redeposición secundaria, por lo que cabe interpretar, al igual que en la Cámara Dinaledi, que la acumulación respondió a un comportamiento funerario. Se han recuperado también algunos restos de fauna.
- Lesedi significa 'luz' en setswana.
- Solo se ha excavado en tres pequeñas áreas de la cámara (UW 102a, 102b y 102c), por lo que es probable el hallazgo de fósiles adicionales en el futuro.
- El descubrimiento de la cámara y de los primeros fósiles (102a) fue realizado por Rick Hunter y Steven Tucker en noviembre de 2013. Los fósiles de 102b y 102c fueron localizados por Hannah Hilbert-Wolf durante el muestreo geológico de la cámara.
- Los fósiles pertenecieron al menos a dos adultos y un menor, si bien se ha propuesto la presencia de un tercer adulto. Están representadas todas las partes esqueléticas, salvo tibia, peroné y pie. Dada la similitud de rasgos, los individuos de Dinaledi y Lesedi podrían pertenecer a la misma población.
- El contexto sedimentario es muy complejo.
 |
| Cámara Dinaledi. Unidades sedimentarias. Dirks et al, 2017. |
La datación directa de los fósiles de la Cámara Dinaledi y sedimentos asociados ha deparado unas fechas entre hace 335-236 ka (Dirks et al, 2017). La supervivencia de Homo naledi hasta una fecha reciente hace muy problemática la clasificación de fósiles y la autoría de artefactos del registro arqueológico atribuidos hasta el momento a H. sapiens arcaicos. La morfología de la mano y la muñeca de H. naledi soportan la hipótesis de que fabricó herramientas. En este caso, habría que revisar el papel de un cerebro grande en la industria lítica.
| Distribución de los restos recuperados en 2013-2014. Bolter et al, 2020. |
 |
| Comparación entre Lee Berger y Homo naledi |
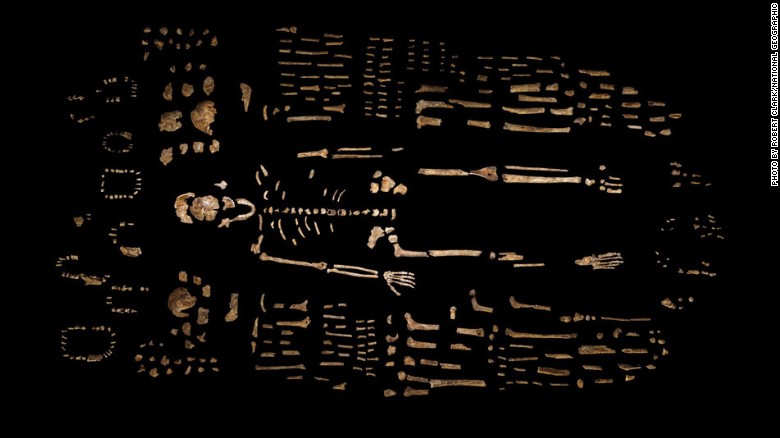 |
| Restos recuperados de Homo naledi en la Cámara Dinaledi |
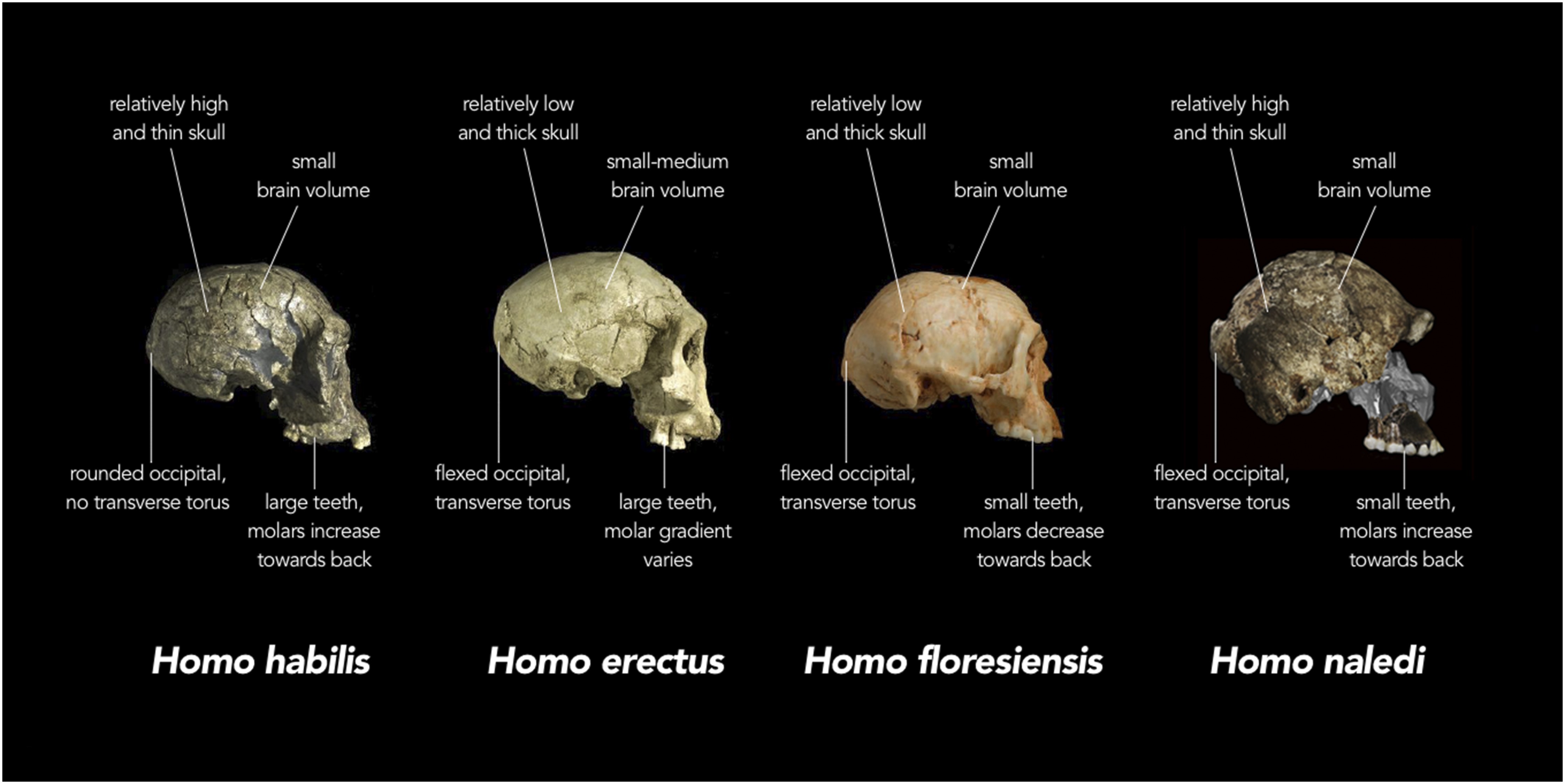
Homo naledi presenta un mosaico de características antiguas y modernas. Se han identificado autapomorfias en el pulgar, la columna vertebral y el fémur proximal (Berger et al, 2015; Kivell et al, 2015; Marchi et al, 2017). El bauplan de H. naledi combina rasgos que no se observan típicamente en especies de Homo (por ejemplo, cerebros y cuerpos pequeños) con otros característicos de H. erectus y especies de Homo más recientes (por ejemplo dimorfismo sexual reducido o miembros inferiores largos). Garvin et al, 2017.
- Cerebro pequeño, de unos 610 cc los varones (LES 1) y 465 cc las hembras, menos de la mitad que el del HAM. La morfología craneal, es similar a la de erectus, habilis o rudolfensis, pero con una combinación de rasgos diferente. Organización del cerebro "moderna" (Holloway et al, 2018). Su cara presenta rasgos arcaicos como un torus supraorbital marcado, cierto prognatismo mandibular y carencia de barbilla (Laird et al, 2016).
- J. Francis Thackeray (2015) ha realizado un análisis morfométrico del cráneo de Homo naledi en base a una regresión lineal de mínimos cuadrados de los valores medios de las mediciones de cráneos de 12 especies de Hominini. De acuerdo con los resultados:
- H. naledi parece ser significativamente diferente a las demás especies.
- H. naledi parece ser más similar a los Homo tempranos, en particular H. habilis, y (en menor medida) H. rudolfensis y Homo erectus.
- Los datos craneales justifican la clasificación de los hallazgos de Rising Star Cave en una nueva especie.
- Un patrón de desarrollo corporal similar al de los humanos o intermedio entre el de los humanos y los hominini tempranos (Bolter et al, 2020).
- Se ha recuperado una mano derecha de adulto completa (falta solo el pisiforme). La muñeca está notablemente derivada y muestra la mayor parte de las características clave que se consideran ventajosas para hacer frente a la alta carga del pulgar durante la manipulación de herramientas. Las articulaciones carpal radial y carpometarcarpal muestran signos de reorientación a una alineación proximodistal más parecida a la humana, incluyendo la forma y expansión del trapecio. Sin embargo, la articulación trapecio primer metacarpal es notablemente pequeña y no existe un proceso estiloides en el tercer metacarpiano. El pulgar es robusto y largo en relación con los dedos, aunque no tanto como en MH2, H. naledi cae dentro de la variación de los varones HAM pero fuera del rango de variación de las hembras. La morfología de la mano sugiere una eficiente capacidad de manipulación y agarre de precisión que solo se han encontrado en neandertales y sapiens. En contraste con la morfología derivada de la muñeca y de la palma, los dedos de H. naledi están fuertemente curvados, más que los de los australopitecinos, incluyendo Au. afarensis, lo que sugiere locomoción arbórea, trepa y suspensión. Por consiguiente, este Homo era capaz de usar sus manos para una manipulación mejorada y al mismo tiempo para la locomoción arbórea (Kivell, 2015).
- El hombro, también es primitivo, conservando adaptaciones a la escalada (Feuerriegel et al, 2016)
- Las vértebras son más similares a las de los miembros del Pleistoceno del género Homo, mientras que la caja torácica es amplia distalmente como en Au. afarensis. Costillas inferiores relativamente planas y robustas. Canal espinal relativamente ancho (Williams et al, 2017).
- Pelvis primitiva. Pala ilíaca semejante a la de Au. afarensis o Au. africanus. Isquion corto con un surco tuberoacetabular estrecho, y ramas isquiopubiana e iliopúbica gruesas, semejantes a Au. sediba y H. erectus. Esta combinación de características ilíaca y isquiopubiana no se ha encontrado en otros Hominini fósiles.
- De tamaño relativamente grande (140-160 cm de altura y 40-55 kg, con escaso dimorfismo sexual), con piernas largas y pies similares a los del HAM, por lo que podría caminar largas distancias. Fémur proximal primitivo.
- La muestra de Dinaledi incluye 107 huesos del pie, incluyendo un pie adulto casi completo.
- El pie del H. naledi es predominantemente moderno, con un hallux aducido, un tarso alargado, y la articulaciones del tobillo y calcaneocuboidea derivadas. Estas características indican un pie bien adaptado para la bipedación.
- Sin embargo, el pie del H. naledi se diferencia del de los humanos modernos en tener más curvas las falanges proximales, por la presencia de un arco longitudinal medial reducido, lo que sugiere un repertorio locomotor único, proporcionando así una prueba más de la diversidad locomotora dentro de los Hominini.
- Fémur, rótula, tibia, y peroné muestran un mosaico rasgos primitivos y derivados e indican adaptación a la marcha a largas distancias (Marchi et al, 2016).
- Dientes pequeños, incluso más que los de los primeros Homo, con morfología oclusal simple. M1<M2<M3. Las coronas molares son modernas, con cinco cúspides, pero las raíces son muy primitivas (Irish et al, 2019).
- El 44% de los dientes muestran daños por traumatismos, una tasa mucho más alta que en otros Hominini. Están más afectados los dientes posteriores que los anteriores, los maxilares que los mandibulares y los de la derecha que los izquierdos. Es poco probable que estos daños se deban a comportamientos culturales no masticatorios. Lo más posible es que los traumatismos fuesen causados por una dieta dura o por contaminantes como la arena. Ian Towle, Joel D. Irish e Isabelle De Groote, 2017. Esto sugiere una dieta muy diferente a la de otros homininos contemporáneos, de forma que no existiría competencia por los recursos (Michael A. Berthaume, Lucas K. Delezene y Kornelius Kupczika, 2018)
- La ontogenia de los dientes deciduos es más similar a la de los humanos que a la de los chimpancés, reflejando probablemente una simplesiomorfia hominini. La ontogenia de los dientes permanentes presenta una mezcla de patrones de los humanos y los chimpancés. Sorprendentemente, el M2 emerge más tarde, como en los humanos modernos, un patrón hasta ahora desconocido en los hominini extintos y que hasta ahora se había considerado que reflejaba una historia de vida lenta. Cofran y Walker, 2017.
- La combinación de número y distribución de los perikymata, líneas de crecimiento visibles en el esmalte dental, es particular para naledi (Guatelli-Steinberg et al, 2018).
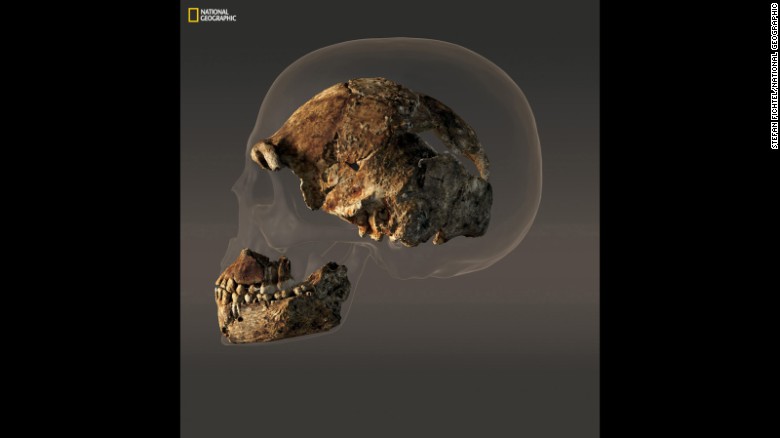 |
| Comparación del cráneo de Homo naledi con el del humano actual. |
 |
| Comparación de rasgos craneales. Hawks et al, 2017. |
 |
| Comparación del cráneo de Homo naledi con el de otros Homo. Chris Stringer, 2015. |
 |
| Comparación de volumen craneal. Hawks et al, 2017. |
 |
| Homo naledi. Reconstrucción. Wits. University. |
 |
| Comparación de anatomía mandibular y dental. Hawks et al, 2017. |
 |
| Homo naledi. DH1. Mandíbula. |
 |
| Comparación del fémur. Hawks et al, 2017. |
 |
| Comparación de rasgos poscraneales. Hawks et al, 2017. |
Holotipo
DH1. Comprende una bóveda craneal parcial, maxilar parcial, y la mandíbula casi completa de un individuo presuntamente masculino.
Las similitudes de H. Naledi con habilis, rudolfensis y erectus, sugieren que aquella tiene sus raíces en el origen inicial y la diversificación de nuestro género, pero nada se puede concretar hasta disponer de dataciones.
Se han propuesto tres escenarios filogenéticos (Berger et al, 2017; ver figura abajo):Otros ejemplares
- DH2. Bóveda craneal parcial que conserva partes del frontal, parietales derecho e izquierdo, temporal derecho y occipital.
- DH3. Bóveda craneal parcial de un individuo presuntamente femenino que conserva partes del frontal, parietal izquierdo, temporal izquierdo, y esfenoides.
- DH4. Bóveda craneal parcial que conserva partes del temporal derecho, parietal derecho y occipital.
- DH5. Bóveda craneal parcial que conserva parte del temporal izquierdo y occipital.
- DH6. Juvenil, a partir de una mandíbula.
- DH7. Esqueleto parcial de un juvenil. Bolter et al, 2020.
- F1. Pie del que falta sólo el cuneiforme medial y las falanges de los dedos II-V.
- H1. Mano derecha de la que solo falta el pisiforme.
- UW 101-377. Fragmento mandibular que conserva la anatomía dental sin desgaste.
- UW 101-484. Diáfisis tibial derecha de la que falta sólo el extremo proximal.
- UW 101-1013. Falange menor con una callosidad indicativa de una fractura incompleta. Thorckmorton et al, 2017.
- UW 101-1142. Mandíbula adulta que presenta un osteoma periférico. Odes et al, 2017.
- UW 101-1391. Fémur derecho proximal que preserva parte de la cabeza, el cuello, parte de los trocánter menor y mayor, y el eje proximal.
- UW 101-1395. Falange menor con una callosidad indicativa de una fractura completa. Thorckmorton et al, 2017.
 |
| LES 1. Barra de escala = 5 cm. Hawks et al, 2017. |
Entre los fósiles recuperados de la Cámara Lesedi, destacan:
- LES 1. Cráneo constituido por 57 fragmentos. Presenta una dentición adulta completa, excepto por las coronas de los incisivos centrales y laterales inferiores izquierdos.
- UW 102a-021. Clavícula derecha completa.
 |
| Homo naledi presenta un mosaico de características modernas y ancestrales. |
Filogenia
- Un origen en algún lugar entre las ramas mal resueltas que conducen a H. habilis , H. rudolfensis , H. floresiensis y Au. sediba (Berger et al, 2015; Dembo et al, 2016; Thackeray, 2015). Esta hipótesis se basa en el análisis de máxima parsimonia de un gran conjunto de datos de rasgos craneales y dentales.
- Gran parte de la evolución de la forma del en Homo parece ser consistente con la evolución neutral por deriva genética, con algunas características que muestran evolución adaptativa (Ackermann y Cheverud, 2004; Weaver et al, 2007; Schroeder et al, 2014). Una rama evolutiva larga crearía una oportunidad sustancial para la deriva. Tal evolución no adaptativa, combinado con la evolución de adaptación de algunos rasgos, podría haber creado el patrón único en esta especie (Laird et al, 2017), aunque parece probable que las características poscraneales estarían sujetos a mayores restricciones adaptativas.
- Una especie hermana de H. erectus y otras especies de cerebro grande, incluyendo H. sapiens (Dembo et al, 2016).
- H. naledi comparte muchas características derivadas de la mano, el pie y los miembros inferiores con H. erectus y H. sapiens que están aparentemente ausentes en H . habilis, H. floresiensis, o Au. sediba. Sin embargo, carece de varios rasgos derivados compartidos por H. erectus y H. sapiens en el hombro, tronco y cadera (Hawks et al, 2017; Williams et al, 2017; Marchi et al, 2017; Feuerriegel et al, 2017).
- En este escenario, el cerebro de gran tamaño en varias especies sería una homología. Además, este escenario requeriría muchos desarrollos paralelos evolutivos en H. naledi y H. sapiens, incluyendo la morfología de la mano y la muñeca (Kivell et al, 2015), el pie (Harcourt-Smith et al, 2015 ), las extremidades inferiores (Marchi et al, 2017), y algunas morfologías craneales y dentales (Laird et al, 2017; Schroeder et al, 2017).
- Dembo et al (2016) aplicaron un marco bayesiano para examinar la posición filogenética de Homo naledi sobre la base de caracteres craneales y dentales, colocándolo como un taxón hermano de un clado que incluye H. antecessor y los Homo posteriores.
- Los rasgos primitivos no compartidos con erectus podrían interpretarse como reversiones evolutivas.
- En este escenario, una población temprana de H. naledi podría haber sido el antepasado temprano de los humanos actuales, situando a erectus en una rama lateral.

Los árboles basados en la morfología craneal indican una estrecha relación entre H. naledi y H. erectus, mientras que el árbol mandibular ubica a H. naledi más cerca del Homo basal. (Schroeder et al, 2016).
También es concebible que las semejanzas craneales y dentales procedan de introgresiones de H. naledi con linajes australolpitecinos durante el Pleistoceno Temprano (Berger et al, 2017).
Enlaces:
- Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. Lee R Berger et al, 2015.
- Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. Paul HGM Dirks et al, 2015.
- W. E. H. Harcourt-Smith, Z. Throckmorton, K. A. Congdon, B. Zipfel, A. S. Deane, M. S. M. Drapeau, S. E. Churchill, L. R. Berger y J. M. DeSilva. 2015. The foot of Homo naledi.
- Tracy L. Kivell, Andrew S. Deane, Matthew W. Tocheri, Caley M. Orr, Peter Schmid, John Hawks, Lee R. Berger y Steven E. Churchill. 2015. The hand of Homo naledi.
- Morphosource. Fósiles en 3D.
- Human evolution: The many mysteries of Homo naledi. Chris Stringer, 2015.
- Homo naledi.org
- Patrick S. Randolph-Quinney. 2015. The mournful ape: Conflating expression and meaning in the mortuary behaviour of Homo naledi
- John Hawks y Lee R. Berger. 2016. The impact of a date for understanding the importance of Homo naledi.
- Elen M. Feuerriegel, David J. Green, Christopher S. Walker, Peter Schmid, John Hawks, Lee R. Berger, Steven E. Churchill. 2016. The upper limb of Homo naledi.
- Myra F. Laird, Lauren Schroeder, Heather M. Garvin, Jill E. Scott, Mana Dembo, Davorka Radovčić, Charles M. Musiba, Rebecca R. Ackermann, Peter Schmid. 2017.The skull of Homo naledi.
- John Hawks, Marina Elliott, Peter Schmid, Steven E Churchill, Darryl J de Ruiter, Eric M Roberts, Hannah Hilbert-Wolf, Heather M Garvin, Scott A Williams, Lucas K Delezene, Elen M Feuerriegel, Patrick Randolph-Quinney, Tracy L Kivell, Myra F Laird, Gaokgatlhe Tawane, Jeremy M DeSilva, Shara E Bailey, Juliet K Brophy, Marc R Meyer, Matthew M Skinner, Matthew W Tocheri, Caroline VanSickle, Christopher S Walker, Timothy L Campbell, Brian Kuhn, Ashley Kruger, Steven Tucker, Alia Gurtov, Nompumelelo Hlophe, Rick Hunter, Hannah Morris, Becca Peixotto, Maropeng Ramalepa, Dirk van Rooyen, Mathabela Tsikoane, Pedro Boshoff, Paul HGM Dirks, Lee R Berger. 2017. New fossil remains of Homo naledi from the Lesedi Chamber, South Africa.
- Lee R Berger, John Hawks, Paul HGM Dirks, Marina Elliott, Eric M Roberts. 2017. Homo naledi and Pleistocene hominin evolution in subequatorial Africa.
- Paul HGM Dirks, Eric M Roberts, Hannah Hilbert-Wolf, Jan D Kramers, John Hawks, Anthony Dosseto, Mathieu Duval, Marina Elliott, Mary Evans Rainer, Grün John Hellstrom, Andy IR Herries, Renaud Joannes-Boyau, Tebogo V Makhubela, Christa J Placzek, Jessie Robbins, Carl Spandler, Jelle Wiersma, Jon Woodhead, Lee R Berger. 2017. The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa.
- Jessica C Thompson. 2017. Human evolution: New opportunities rising
Te puede interesar:
|
No hay comentarios:
Publicar un comentario