2,2–1 Ma. En base al descubrimiento en Kromdraai de TM 1517, el único esqueleto parcial de esta especie conocido hasta ahora (Broom, 1938a, Broom 1938b, Broom 1942 y Broom, 1943). Estos hallazgos fueron también los que motivaron la propuesta del género Paranthropus por Broom en 1938. El escolar Gert Terblanche localizó en 1938 los primeros restos: un cráneo parcial y mandíbula.
Fragmentos de cráneo incluyendo cinco dientes y pocos fragmentos de esqueleto.
Único esqueleto parcial de esta especie conocido hasta la fecha.
Las circunstancias del hallazgo y el contexto estratigráfico son confusos.
Algunos de los restos que componen TM 1517, se considera actualmente que pertenecieron a cercopitécidos.
La desaparición de la especie pudo estar relacionada con un cambio climático (Caley et al, 2018).
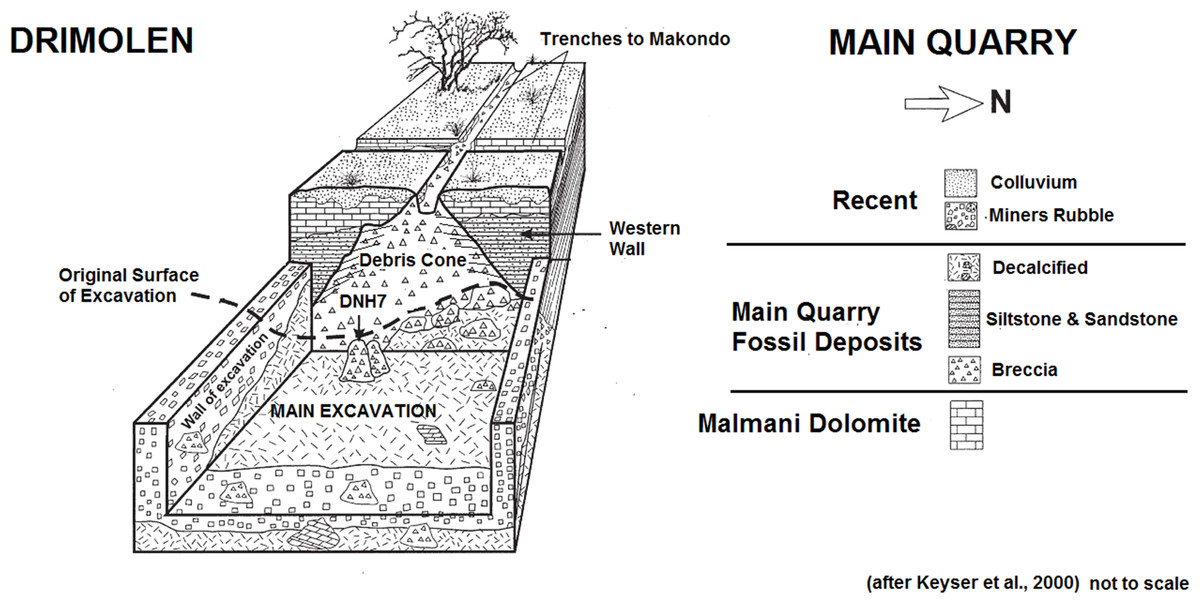
Kromdraai B (KB) (Gauteng, Sudáfrica). 2-1,5 Ma. Es un vano de dolomita rellenado de depósitos fosilíferos, 2 km al este de Sterkfontein. La datación es imprecisa y, frente a la hipótesis más aceptada de que Paranthropus tiene su origen en Africa del Este con P. aethiopicus, se ha sugerido que al menos algunos especímenes de Kromdraai datan del origen del género Paranthropus (Kaszycka 2002 y Tobias 1988) y son intermedios entre los hominini de Makapansgat Miembros 3/4 y Sterkfontein Miembro 4, por un lado y los más derivados de Swartkrans por otro. Las excavaciones fueron dirigidas por Broom (1938-1944), Brain (1955-1956), Vrba (1977-1980) y Thackeray (1993-2002). Desde 2002, las excavaciones corren a cargo del Kromdraai Research Project. Los contextos para las excavaciones de Broom y Brain son confusos, imposibilitando una datación precisa. Los carnívoros fueron el principal agente de acumulación.
Individuo 1. Adolescente tardío. TM 1517. Broom (1938a); Broom y Schepers (1946); Broom (1943); Broom (1938b); Day y Thornton (1986); Day (1978); Skinner et al. (2013); Broom (1942).
Individuo 2. Juvenil. Edad al morir, 1,63-2,02 años (Smith et al, 2015). TM 1536. Fragmento mandibular izquierdo con I1, I2, di2, dm1-2, M1; L dC. Broom (1941); Broom y Schepers (1946).
Individuo 3. Adulto. TM 1600. Dos fragmentos del corpus mandibular, con M2-M3 (fragmento1) y P3 (fragmento 2). Brain (1981).
Individuo 14. Adulto. KB 5383. M1 superior derecho. Vrba (1981).
Individuo 15. Adulto. KB 5389. I1 superior izquierdo. de Ruiter (2004).
Individuo 16. Juvenil. KB 5503. dm2 inferior derecho.Thackeray et al (2001).
Posiblemente Individuo 1. KB 5522. Fragmento del eje del húmero izquierdo. Thackeray et al (2005).
Posiblemente Individuo 1. KB 5524. Cara lingual de una corona molar desgastada, posiblemente M1 o M2.Braga et al (2013).
Individuo 17. Juvenil. KB 6067 es una porción petrosa del hueso temporal de un individuo juvenil. Su morfología lo relaciona con P. robustus, pero también con StW 53 (Homo gautengensis). Sin embargo, el tamaño de la ventana oval es más parecido al de la de algunos especímenes de Sterkfontein Member 4 (Australpithecus africanus). Si KB 6067 se incluye dentro de P. robustus, esto puede representar una condición evolutivamente menos derivada que la deducida a partir de TM 1517 y otros ejemplares conespecíficos. Braga et al (2013).
Kromdraai E (KE). Excavaciones a partir de 2014 a cargo del Kromdraai Research Project. Hallazgos pendientes de publicación.
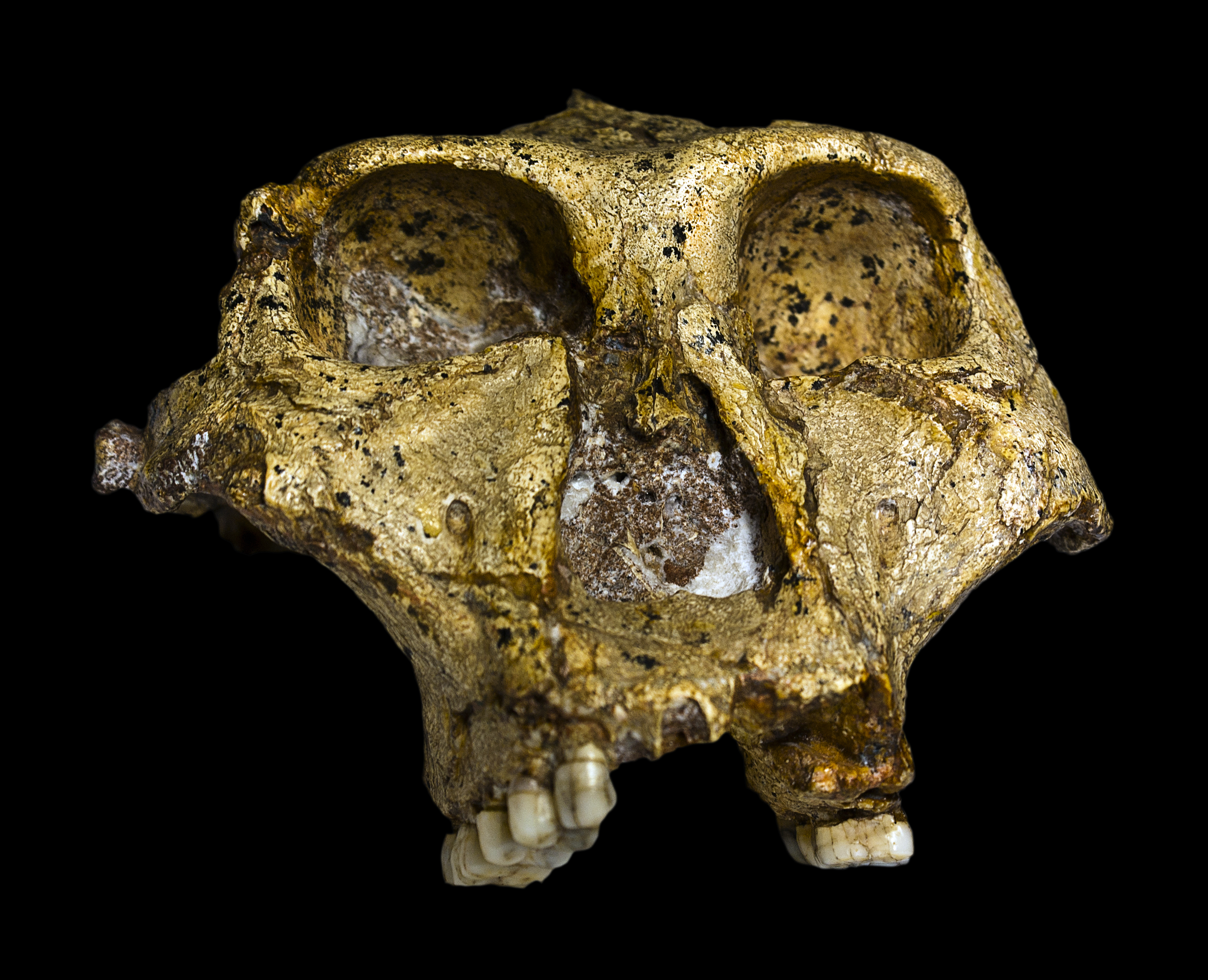
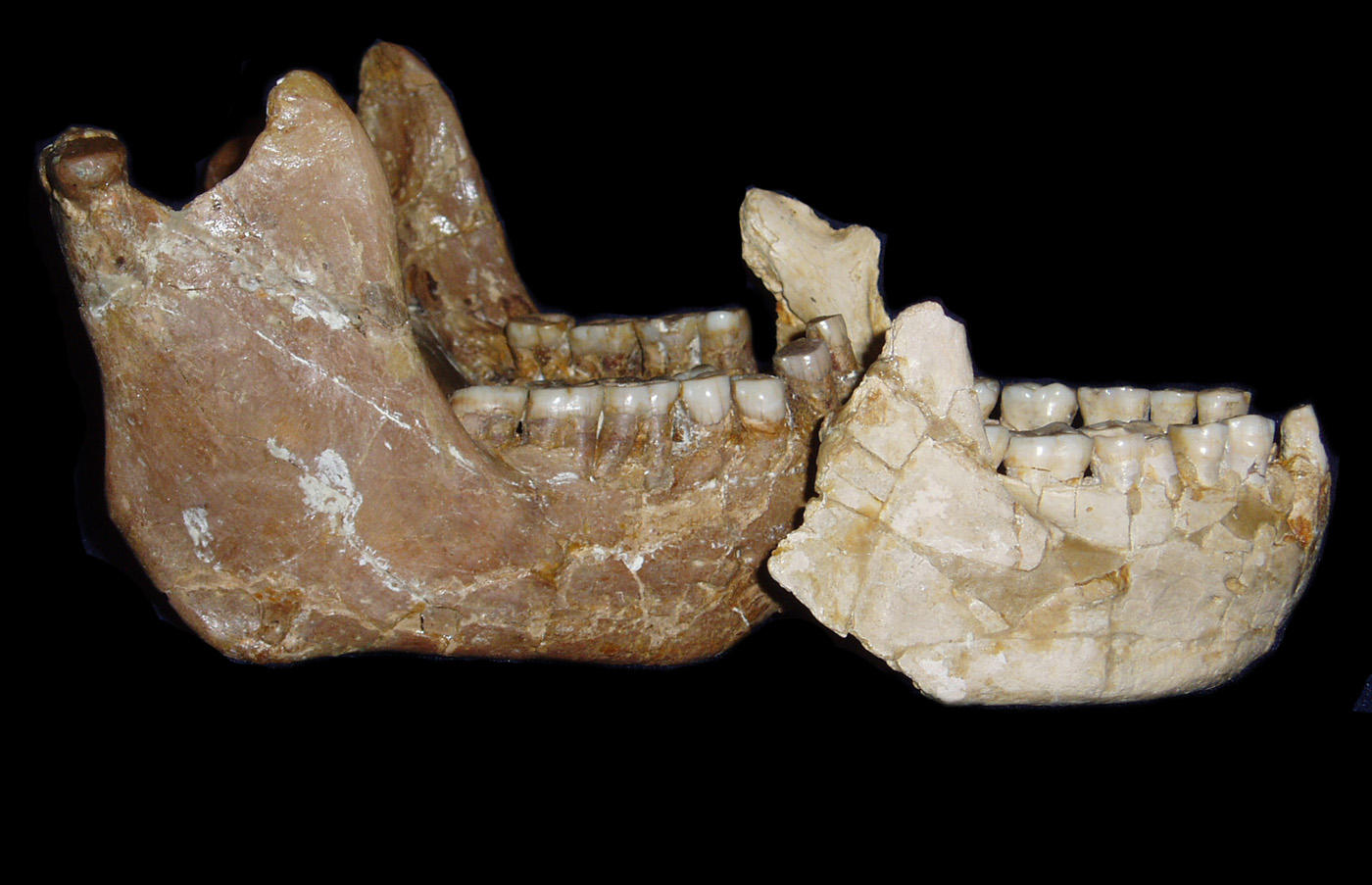
Swartkrans. Miembro 1 (2,25-1,7 Ma), Miembro 2 (1,7 Ma) y Miembro 3 (1 Ma) (Pickering et al, 2011; Gibbon et al, 2014). Es el yacimiento con una muestra más amplia de Paranthropus robustus. Los ejemplares de Swartkrans muestran unos dientes mucho más grandes y por ello se ha sugerido que representan una especie diferenciada: Paranthropus crassidens (Broom 1949, Broom 1950, Howell 1978, Grine 1982, Grine 1985 y Grine 1988). Sin embargo, los restos dentales de Drimolen, de tamaño intermedio, favorecen la hipótesis de una única especie (Keyser et al, 2000 y Moggi-Cecchi et al, 2010).
SK 12. Cráneo. Macho.
SK 46. Cráneo. Macho.
SK 48. 1950. Mr. Fourie. Se trata de un cráneo, de hembra adulta (macho para Charles A. Lockwood et al (2007). Es
el cráneo más completo de los robustus.
Estatura 1,32-1,10,
un poco superior a la de los australopithecus.
Peso 40-32 kg.
Bipedación casi
humana.
Los restos de las
manos indican mayor finura táctil que la de los australopitecus.
Según Sandi Copeland (2011), tras estudiar la procedencia de
los individuos a partir de rastros en los dientes, los grupos de africanus practicaban la filopatría (los
machos se unían a hembras de fuera).
Para Charles A. Lockwood et al (2007)P. robustus muestra un alto dimorfismo sexual y un desarrollo retardado de los machos con respecto a las hembras. Este patrón sugiere que la estrategia reproductora masculino consistía en monopolizar grupos de hembras, de una manera similar a como lo hacen los gorilas de espalda plateada. El costo para los varones debió ser una alta tasa de depredación, lo que justifica la abundancia de machos jóvenes en el registro fósil.
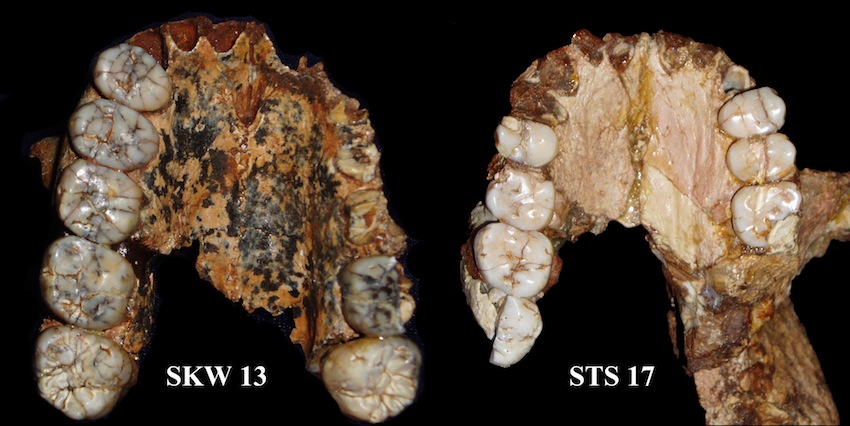
SKW 13 Paranthropus robustus y STS 17 Australopithecus africanus
Características
Grandes y anchos molares y premolares. Caninos e incisivos pequeños.
Mandibulas masivas, cortas, altas y extremadamente robustas con forma de U divergente (mastiación potente). No tanto como boisei. Corpus amplio y profundo y una alta rama, relativamente vertical. No se descarta que, aparte de la dieta vegetal, ingiriese insectos. Según los estudios de Andrew Sillen y Julia Lee-Thorp, P. robustus ingería proteínas animales.
Caja craneal bastante alta y desarrollada, con cresta sagital.
Volumen cerebral 530 cc.
Enorme hueso malar.
Rostro recto, con arco cigomático muy ancho y robusto. Torus supraorbitario masivo.
Esmalte dental muy grueso, mucho más que el A. africanus. Según Wynn, Cerling y Sponheimer (2013), su dieta estaba compuesta de alimentos procedentes de árboles y arbustos, pero también de hierbas y juncias, con gran variación entre los individuos. Según Ungar y Hlusko (2016), a pesar de su aparatosa dentadura, solo consumía alimentos duros ocasionalmente.
Los australopitecinos robustos del Este de África también tienen esmalte muy grueso en los molares.
Podría ser una sinapomorfia, lo que implicaría una rama robusta común antes de la separación.
Podría ser un rasgo desarrollado por separado en cada especie, lo que convertiría en dudoso el clado robusto.
Tamaño y proporciones semejantes a los Australopithecus pero más robustos.
Broom, Robert, and John Talbot Robinson. "Man contemporaneous with the Sawartkrans ape‐man." American journal of physical anthropology 8.2 (1950): 151-156.
Robinson, John Talbot. "Further remarks on the relationship between “Meganthropus” and australopithecines." American journal of physical anthropology 13.3 (1955): 429-445.
Sillen, Andrew. "Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus and associated fauna from Swartkrans." Journal of Human Evolution 23.6 (1992): 495-516.
Lee-Thorp, Julia A., Nikolaas J. van der Merwe, and C. K. Brain. "Diet of Australopithecus robustus at Swartkrans from stable carbon isotopic analysis." Journal of Human Evolution 27.4 (1994): 361-372.
Keyser, André W. "The Drimolen skull: the most complete australopithecine cranium and mandible to date." South African Journal of Science 96.4 (2000): 189-192.
Blackwell, Lucinda, and Francesco d'Errico. "Early hominid bone tools from Drimolen, South Africa." Journal of Archaeological Science 35.11 (2008): 2880-2894.
Moggi-Cecchi, Jacopo, et al. "Early hominin dental remains from the Plio-Pleistocene site of Drimolen, South Africa." Journal of Human Evolution 58.5 (2010): 374-405.
DNH 134, un fragmento de cráneo, y KNM-ER 2598, un fragmento occipital, son los únicos ejemplares africano de erectus más antiguos que los fósiles de Homo de Dmanisi y Java. Se han datado en hace 2,04-1,95 Ma y 1,87 Ma respectivamente.
Los erectus más antiguos, provienen de Koobi Fora, Kenia, y la especie persiste en África hasta cerca de la frontera Brunhes-Matuyama (0,78 Ma).
Los restos son más abundantes en Kenia y Tanzania, aunque también se han hallado ejemplares en Etiopía y en el Sur.
Los ejemplares mejor conservados se hallaron en las riberas del lago Turkana. Se complementan
con los hallazgos de la garganta de Olduvai.
Tobias y von Koegniswald
(1964) definieron los grados de hominización:
Australopitecos.
Habilis.
Erectus de Sangiran y
ejemplares de Olduvai como OH13.
OH13 había sido considerado por L.
Leakey (1964) como paratipo del habilis.
Esto indica bien a las claras las dificultades con que tropezamos a
la hora de establecer fronteras precisas en la transición entre habilis y erectus.
Erectus de Olduvai,
Ternifini, Kabuh y Zhoukoudian.
El fragmento de cráneo DNH 134, datado entre hace 2,04-1,95 Ma, hallado en Drimolen, África del Sur, presenta similitudes con el cráneo de Sangiran (Herries et al, 2020).
Un par de cráneos de
Turkana presentan rasgos similares al erectus
de Pekín, pero con una capacidad craneal inferior.
KNM-ER 3733 (Leakey y Walker, 1985) fue descubierto por Bernard Ngeneo, del equipo de Richard Leakey, en 1975 en Koobi Fora (Kenia). Se trata de un cráneo completo de hembra madura, con capacidad craneal de 850 cc, similar a la del Hombre de Pequín. Se ha datado en ~1.63 Ma (Lepra y Kent, 2015).
KNM-ER 3883 (Leakey y Walker, 1985). Cráneo de 800 cc datado en 1,6 Ma.
Del mismo yacimiento es KNM-ER 1808
(esqueleto parcial).
En el yacimiento etíope de Gona, situado en el triángulo de Afar, se recuperaron herramientas líticas sencillas, de tipo Olduvayense (modo 1), junto con otras más complejas, de tipo Achelense (modo 2), asociadas a restos de erectus (Semaw et al, 2020):
DAN-5: Un cráneo casi completo recuperado en la zona norte del río Dana Aoule cuya antigüedad se estima en 1,5 Ma. Con un volumen endocraneal de 590 cc es el de menor volumen de los erectus africanos conocidos. Muestra similitudes con los ejemplares de Dmanisi.
BSN-12: Un cráneo incompleto de hace 1,26 Ma descubierto en el norte del río Busidima (BSN-12) a 5,7 kilómetros de distancia del anterior.
El cráneo BSN-12 es grande y resistente (similar al OH9 de la Garganta de Olduvai, Tanzania), mientras que DAN-5 es más pequeño y flexible, lo que sugiere que erectus mostraba acusado dimorfismo sexual.
El equipo de Domínguez-Rodrigo
halló en verano de 2009, en el yacimiento de Olduvai un fémur y un radio de
adulto, posiblemente del mismo individuo, datados en 1,3 Ma. La cabeza del
radio es un tercio más grande que la del sapiens
y la tuberosidad donde se inserta el bíceps duplica a la del hombre actual.
Grado erectus finales
El norte del Danakil (Afar, Eritrea) ha proporcionado desde mediados de los noventa evidencia sobre el erectus, con el descubrimiento de más de 200 sitios de finales del Pleistoceno en un afloramiento sedimentario fluviolacustre de 1.000 m de espesor en la cuenca del río Dandiero, al sur de Buia (Abbate et al, 1998). Las excavaciones corren a cargo de la Expedición Danakil Italiana y Eritrea, coordinada por la Universidad La Sapienza de Roma y el Museo Nacional de Eritrea, bajo la dirección de Alfredo Coppa, dentro del Proyecto Internacional Buia.
El yacimiento de Uadi Aalad (UA), consta de depósitos deltaicos y fluviales de 5-6 m de espesor radiométricamente y biocronológicamente fechados en ca 1 Ma (Ghinassi et al, 2009). Destacan los hallazgos en la capa LK3 Homo (Macchiarelli et al 2004; Bondioli et al, 2006).
UA 31. Señora de Buia. Descubierto por el equipo de Lorenzo Rook y Berhane Tesfamariam. Un cráneo de adulto prácticamente completo con la cara preservada, atribuido a hembra, con mezcla de características erectus/ergaster y rasgos derivados, incluyendo caja craneana larga y ovalada, de más de 20 cm de longitud, un alto posicionamiento de los puntos más externos parietales como en rhodesiensis, una angulación débil a lo largo de la línea media y un ligero aplanamiento parasagital y huesos parietales delgados. No se ha determinado si su prominencia parietal es una consecuencia estructural de dolicocefalia, la expresión de la variación individual/regional, o una característica derivada con posible relevancia filogenética. Acusados arcos supraorbitales.
Según Bruner et al (2016), su capacidad craneal es de 995 cc. Las áreas occipitales muestran una pronunciada abultada, el cerebelo se encuentra en una posición posterior, y los vasos meníngeos medios están más desarrollados en las regiones posteriores. Estas características son comunes entre los especímenes atribuidos a erectus, en particular los endomoldes del Pleistoceno Medio de Zhoukoudian. Los lóbulos parietales muestran una curvatura pronunciada, asociada a una base del cráneo estrecha.
UA 222 y UA 369. Dientes incisivos permanentes. Tienen un esmalte relativamente delgado en comparación con la condición humana moderna, como en Homo antecessor, y la morfología recuerda al heidelbergensis norteafricano.
UA 173, UA 405 y UA 466. Fragmentos de pelvis.
En Mulhuli-Amo (MA), a 4,7 km de Uadi Aalad y formando parte posiblemente del mismo horizonte estratigráfico se han hallado nueve fragmentos humanos, pertenecientes posiblemente a tres individuos.
MA 14 fragmento de frontal.
MA 64 y MA 88 a-f, fragmentos parietales y MA 89 fragmento temporal, pertenecientes al mismo individuo adulto.
MA-88, compuesto por los seis fragmentos a-f, constituye un parietal izquierdo casi completo, incluyendo la región del asterión, cuyo espesor y características arquitectónicas y estructurales se ajustan con precisión a UA 31.
MA 93 Corona sin uso de un molar permanente inferior.
La campaña de 2016 ha proporcionado huellas de erectus datadas en hace 800 ka, en los sedimentos fluviales de un paleolago rodeado de pastizales, en el yacimiento de Aalad-Amo.
Daka. BOU-VP-2/66
Daka (o Dakanihylo, Formación Bouri, Middle Awash, Etiopía) Asfaw et al, 2002. BOU-VP-2/66. Caja craneana descubierta por Henry Gilbert, del equipo de Berhane Asfaw y Tim White, en 1997, datada en 1 Ma y con una forma similar a la de Buia. Según Keely B. Carlson et al (2013) es similar en morfología lineal a los fósiles de los erectus asiáticos. Para Baab (2016), pudo haber pertenecido a una población "avanzada" de erectus cercana a la raíz del Homo heidelbergensissensu lato, o a una población inicial de H. heidelbergensis sensu lato.
Olorgesailie. KNM-OG 45500. Cráneo parcial datado en 0,9 Ma, descubierto en 2003 por el equipo de Rick Potts. Capacidad craneal de unos 700-800 cc. Se recuperó aproximadamente 1,5 km al este de una gran acumulación de hachas de mano, que provienen de la misma capa.
Tighenif 1 y 3
Tighenif 1 y 3
Otros hallazgos
MK3. En las excavaciones dirigidas por J. Chavaillon en 1976 en Gombore I (Melka Kunture, Etiopía) apareció in situ, en la unidad 2 del nivel 3, datada en >1,39 Ma, una porción distal bien conservada de un húmero izquierdo de hominini conocido como Gombore IB-7594, Melka Kunture 3 o MK3, asociado a un rico conjunto lítico de transición olduvaiense achelense. De acuerdo con el análisis deFabio Di Vincenzo et al (2015), MK3 añade una gran cantidad de variabilidad al género Homo. El gran tamaño de MK3 sugiere un peso corporal cercano a 90 kg, lejos de la gama del tamaño del cuerpo conocido para los Homo del Pleistoceno temprano.
Los autores sugieren que la dimensión y la morfología de MK3 pueden ser considerados como una exaptación que se convirtió en útil cuando los primeros humanos poblaron altas altitudes, como la cuenca superior del Awash, en la meseta etíope, por encima de 2.000 m de altura. El húmero fue estudiado también por Lague (2015).
OH 9, Chellean Man (Heberer, 1963; Rightmire, 1979). Descubierto por Louis Leakey en 1960 en la garganta de
Olduvai (Tanzania). Su edad se estima en 1,5 Ma. Consiste en una bóveda craneal
de unos 1065 c.c. de capacidad.
OH 12, Pinhead (Rightmire, 1979). Descubierto
por Margaret Cropper en 1962 en la garganta de Olduvai (Tanzania) Es parecido, pero
menos completo que OH 9, y su capacidad craneana es menor, sólo 750 c.c. Muy
similar a KNM-ER 3733, aunque un millón de años más joven. Su edad está comprendida
entre 600 y 800 ka.
KNM-ER 730 (Wood, 1991). Occipital, parietal, frontal y mandíbula parciales.
KNM-ER 736 (Rightmire, 1990). Fémur.
KNM-ER 737 (Rightmire, 1990). Fémur.
KNM-ER 820 (Wood, 1991). Mandíbula de subadulto.
KNM-ER 992 (Wood, 1991). Mandíbula. Holotipo de Homo ergaster.
KNM-ER 1472. (Wood y Collard, 1999). Fémur.
KNM-ER 1481. (Wood y Collard, 1999). Fémur.
KNM-ER 1808 (Walker, Zimmereman y Leakey, 1982). Esqueleto y fragmentos craneales.
KNM-ER 3228. Coxal.
KNM-ER 3733 (Leakey y Walker, 1985). Cráneo.
KNM-ER 3883 (Leakey y Walker, 1985). Cráneo.
KNM-ER 42700 (Spoor et al, 2007). Calvaria de adulto joven hallada en Ileret, Kenia, datada en 1,55 Ma. Su atribución taxonómica está sujeta a controversia (Baab, 2016). El tamaño es de los más pequeños conocidos para erectus. Los análisis morfogeométricos dan como resultado una forma intermedia entre erectus y el HAM (Catherine C. Bauer y Katerina Harvati, 2015).
KNM-WT
15000, Turkana Boy Descubierto por Kamoya
Kimeu en 1984 en la ribera del río Nariokotome cerca del lago Turkana (Kenia). (Brown et al. 1985; Leakey and Lewin 1992;
Walker and Leakey 1993; Walker and Shipman 1996). Se trata
de un esqueleto casi completo de un niño entre 7,6-8,8 años, según estimación a partir de los parámetros dentales (Dean y Smith, 2009) al que le faltan las
manos y pies. Es el Homo erectus más completo conocido, con una edad de 1,5 Ma. Su capacidad craneal es de 880 cc, y hubiera alcanzado los 910 cc de completar su desarrollo. Su
talla era de 1,60 m. Christopher B. Ruff y M. Loring Burgess (2014) han estimado una estatura adulta de 176-180 cm y una masa corporal adulta de 80-83 kg en base a un patrón de crecimiento similar al de los simios africanos. Los músculos debían de estar muy
desarrollados. Excepto por el cráneo, el esqueleto es muy similar al de los niños
actuales, con pequeñas diferencias. La forma y el tamaño del canal espinal son similares a los de los HAM (Meyer y Haeusler, 2015). Se ha relacionado con diferentes patologías, sin que exista una conclusión definitiva. La maduración dental y esquelética es más parecida a la de un simio que a la de un humano moderno. El aparato locomotor muestra características totalmente modernas, con posibles adaptaciones para la carrera de resistencia (Marchi et al, 2019).
KNM-WT 51260, tercer metacarpiano hallado en Kaito (oeste del Lago Turkana) en 2012, datado en 1,4 Ma. Según el estudio de Carol WARD V. et al (2013) el hueso se asemeja al de un humano moderno en proporciones generales y morfología. Es el tercero más largo de los Hominini conocidos anteriores a los neandertales y humanos modernos tempranos. En particular, muestra una apófisis estiloides bien desarrollada, una característica distintivas de la mano moderna y neandertal Neandertal, no presente en los primeros Hominini. Esta proyección ayuda a estabilizar la muñeca cuando la mano está agarrando pequeños objetos entre el pulgar y los dedos. La morfolgía y función de la mano modernas estaban presentes dentro del contexto de la tecnología achelense y sugiere que las actuales características carpometacarpianas evolucionaron temprano, seleccionadas para un mejor manejo de las herramientas.
BSN49/P27. Gona (Simpson et al, 2008). Pelvis. Debió pertenecer a una hembra de tamaño pequeño, pero con una cadera amplia para ese tamaño.
SK 15 (Robinson, 1961; Howell, 1978; Grine, 2001). Mandíbula parcial.
SK 45 (Robinson, 1961; Howell, 1978; Grine, 2001). Mandíbula parcial.
SK 847 (Clarke, Howell y Brain, 1970; Tobias, 1991; Walker, 1981; Kimbel, Johanson y Rak, 1997; Grine, 2001). Restos mandibulares, dentales y cráneo parcial, datado en hace 2,3-1,65 Ma. Hay daño provocado por un absceso, y probablemente al menos otros dos, en la cara anterior del maxilar y asociados con los incisivos (Ian Towle y Joel D. Irish, 2019).
SK 1896 (Susman, de Ruiter y Brain, 2001). Fragmento distal de fémur.
SK 2045 (Susman, de Ruiter y Brain, 2001). Fragmento proximal de radio.
SKW(SKX) 34805 (Susman, de Ruiter y Brain, 2001). Fragmento distal grande de húmero.
GAR IVE (Condemi, 2004; Zanolli et al, 2016). Mandíbula parcial inmadura hallada en Garba, Melka Kunture, en 1981. Se trata del representante humano más antiguo en un entorno montañoso, datado en hace ca 1,7 Ma.
Atlanthropus mauritanicus
En 1954-1955 C. Arambourg y R. Hoffstetter hallaron en Tighenif, a 20 km de Mascara, Argelia varios restos incluyendo las mandíbulas Tighenif 1 y 3.
Con los hallazgos, Arambourg creó un nuevo género y especie aunque posteriormente se han clasificado dentro de Homo erectus (e.g., Howell, 1960; Geraads et al., 1986; Rightmire, 1990). También podría tratarse de un heidelbergensis muy temprano.
¿heidelbergensis africano?
Los conocimientos actuales sugieren una dicontinuidad filogenética en el momento de la reversión Matuyama / Brunhes hace 780 ka, en posible relación con el fenómeno más general conocido como la Revolución del Pleistoceno Medio (Maslin y Ridgwell, 2005) que, a su vez, corresponde con los drásticos cambios climáticos de MIS 18-16. El periodo entre 900-600 ka es muy pobre en fósiles, pero parece que una especie más encefalizada surge en África y se extiende rápidamente por África y Eurasia. (Homo heidelbergesis según Rightmire, 1998, 2008; Mounier et al, 2009, 2011; Stringer, 2012; pero Arsuaga et al, 2014, 2015; Balter, 2014) donde se encuentra con otros Homo (Profico, 2015).
En la capa de achelense de Gombore II, en la zona arqueológica Melka Kunture, el alto valle de Awash, región de Oromia, a unos 50 km al sur de Addis Abeba, a más de 2.000 msnm, aparecieron dos grandes fragmentos craneales, con una datación de ca 850 ka.
En 1973, un parietal izquierdo parcial (Melka Kunture 1, MK1; Oakley et al, 1977), clasificado como Homo cf. erectus (Chavaillon et al 1974; Chavaillon y Coppens, 1975, 1986)
En 1975, una parte derecha del frontal (Melka Kunture 2, MK2).
Probablemente pertenezcan al mismo cráneo.
Con una datación de hace 700 ka se descubrieron huellas humanas, de otros mamíferos y aves, junto a un hipopótamo despiezado. Las huellas descubiertas pertenecen tanto a adultos como a niños, algunos de los cuáles podrían tener un año de edad. Nos sugieren una infancia con más responsabilidades y menos supervisión adulta que en la actualidad (Altamura et al, 2018).
Recreación de la escena según las huellas, herramientas y fauna -Matthew Bennett-
Junto a las huellas de los seres humanos aparecieron los restos de un hipopótamo que había sido despiezado por los miembros del grupo, además de escamas de piedra y herramientas confeccionadas allí mismo para realizar la tarea de la obtención de la carne.
Huella del pie izquierdo de un adulto - Matthew Bennet -
Dentro del conjunto de herramientas se aprecian todas las secuencias de reducción lítica, lo que sugiere que fueron talladas en el mismo lugar del procesado de la carne del hipopótamo. El material utilizado fue la obsidiana; a nivel tipológico los restos líticos se adscriben al Achelense Medio.
El descubrimiento de las huellas de los niños revela que los pequeños acompañaban a los adultos a realizar las tareas más importantes para su vida cotidiana y supervivencia, probablemente con la intención de que aprendieran in situ las tareas de talla de herramientas y despiece de los animales desde edades muy tempranas.
Tampoco se descarta que las huellas puedan evidenciar juego y entretenimiento de los pequeños mientras los adultos completaban el trabajo. Lo único seguro que podemos concluir es que los niños acompañarían a los grupos móviles de caza, adentrándose en situaciones peligrosas y en la que los adultos no ejercerían la sobreprotección de los hijos a la que estamos acostumbrados en la actualidad.
Atendiendo a la morfología de los seres humanos aparecidos en cronologías similares en Melka Kunture, se supone que las huellas pertenecerían a miembros de la especie Homo Heidelbergensis.
Para Profico et al (2015) MK1 y MK2 representan un único cráneo de morfología arcaica en su curvatura sagital y transversal:
Rasgos comunes con otros ejemplares arcaicos: ausencia de foramen parietales, el desarrollo de la rama media de los vasos meníngeos, las líneas temporales que recorren el parietal medialmente a la eminencia pariental, las marcadas líneas temporales sobre el hueso frontal y la presencia de un pesado toro frontal.
Rasgos peculiares: espesor notable y fuerte divergencia de las líneas temporales tras la constricción postorbital.
El cráneo muestra afinidades con los erectus africanos (ergaster) y heidelbergesis. MK1 está más cerca de erectus en la curvatura y forma del perfil sagital medio y más cerca de heidelbergensis en las dimesiones absolutas y la curvatura y la forma generales.
Tras la reconstrucción digital, la capacidad craneal se ha estimado en ca 1.080 cc.
El cráneo perteneció a un individuo de unos 35-40 años en el momento de su muerte.
De acuerdo con el análisis, estos ejemplares llenan un vacío fenético entre grado erectus y heidelbergensis y representan el mejor candidato o el único para el surgimiento de Homo heidelbergensis alrededor de hace ca 800 ka, así como una evidencia de que esta especie probablemente se originó en África antes de su dispersión en Eurasia.
Datación de los principales yacimientos de Eurasia correspondientes al Paleolítico Inferior y eventos climáticos.
Pocas respuestas
Los hallazgos de Dmanisi (Homo georgicus) marcan el inicio de la ocupación humana de Europa. Las evidencias más antiguas de esta ocupación están representadas por:
Kozarnika, Bulgaria (1,6-1,4 Ma. Sirakov et al, 2010). Molar humano, industria de núcleo y lascas no achelense y huesos incisos que pueden ser el resultado de un comportamiento simbólico.
Pirro Nord, Italia (1,6-1,3 Ma. Arzarello y Peretto, 2010). Hallazgos líticos.
Atapuerca y Orce (España).
La presencia humana parece limitada al sur de la latitud 45ºN, constituida por grupos con tecnologías Modo 1 (Jöris, 2013).
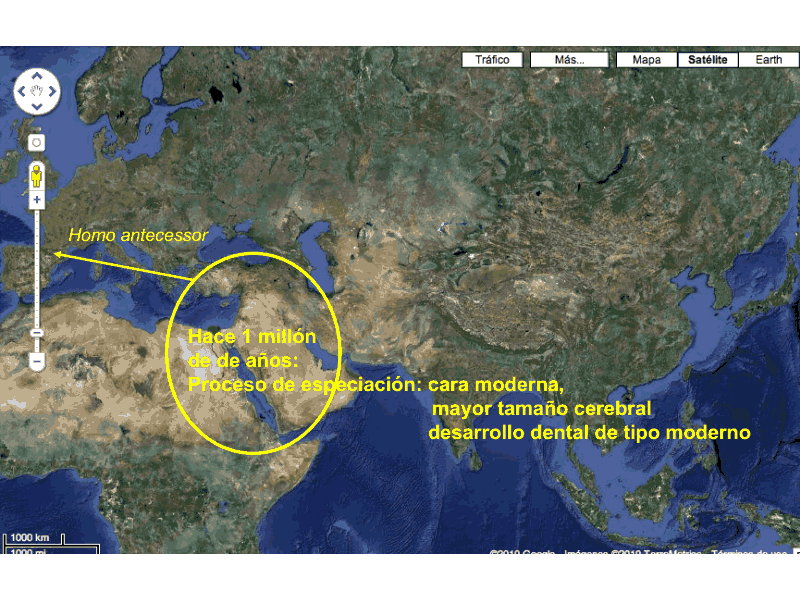
Los restos de Atapuerca, datados en 1,3-0,73 Ma demuestran una ocupación temprana de Europa. Quienes eran aquellos Homo y por dónde llegaron, son preguntas difíciles de responder. Quizá estuvieron en Europa algunos contingentes de erectus. Se han comunicado algunos hallazgos, escasos y controvertidos. Si lo hicieron, se extinguieron pronto o constituyeron grupos marginales. Los hombres de la Sima del Elefante se diferencian del erectus, pero guardan algunas similitudes con los ejemplares hallados en China. Es posible que procedieran de la misma migración de África hace 1,9 Ma. Su viaje por Europa debió recorrer el Corredor de Levante, al mismo tiempo que sus organismos se adaptaban a las condiciones ambientales hasta constituir un demo diferenciado que daría lugar al Homo antecessor.
Yacimientos del Paleolítico Inferior en Eurasia Occidental y Mediterráneo. Los círculos completos indican yacimientos líticos y los semicírculos yacimientos líticos y con restos humanos.
La especie ñ
Los primeros fósiles europeos se datan entre 1,4 y 0,9 Ma. No ha sido identificada la especie a la que pertenecen. El equipo de Atapuerca considera que se trata de un nuevo taxón al que denominan provisionalmente "especie ñ", debido a que la mayor parte de los hallazgos corresponden al territorio español.
HOMO EN EUROPA ENTRE 1,3 Y 0,73 MA
PERIODO MA
FÓSILES
TECNOLOGÍA
1,4-0,9
No ha sido identificada la especie, que se denomina, provisionalmente, especie ñ
Modo 1 muy pobre
0,9-0,73
Homo antecessor
Modo 1 evolucionado
0,7-0,615
Homo antecessor en proceso de extinción.
Oleadas migratorias sin éxito demográfico.
Modo 1
Modo 2 pobre
0,615-0,2
Grado Heidelbergensis
Modo 2 pleno
0,2-0,027
Homo neanderthalensis
Modo 2
Modo 3. Musteriense.
Estratigrafía de los yacimientos ibéricos anteriores al subcrón Jaramillo. Joan García et al, 2014.
Hombre de Orce
Descubierto en 1982 en las excavaciones dirigidas por J.
Gibert Clols en la cuenca de Baza (Venta Micena, Orce, Granada). Durante el
Pleistoceno Inferior existió allí un gran lago interior que desapareció hace 100 ka. No había cuevas y la vida de los homínidos se desarrollaba al aire libre. Gracias a las aguas termales sublacustres, a unos 36 ºC, el lago mantuvo un nivel freático relativamente permanente, con temperaturas más cálidas durante todo el año. Esto dio como resultado un alto nivel de productividad orgánica, especialmente en el Calabriense, lo que permitió el desarrollo de una comunidad de mamíferos rica y bien diversificada, similar a la de las sabanas africanas modernas. La alta biomasa de ungulados favoreció a los carroñeros, como las hienas y los homínidos, que dispusieron de abundantes canales, lo que explica las altas densidades de restos óseos conservados en los sedimentos distribuidos a lo largo de los alrededores del lago (José Manuel García-Aguilar et al, 2014). El análisis de microvertebrados de las unidades D1 y D2 de Barranco León revela que las temperaturas eran significativamente más altas que 13ºC, con condiciones húmedas prevalentes (Agustí et al, 2015).
El triángulo comprendido entre Orce y sus dos pedanías, Fuente Nueva y Venta Micena ofrece la mayor densidad de huesos fósiles de mamíferos cuaternarios conocida hasta el momento en Eurasia. La ausencia de suidos en las capas asociadas a restos humanos permiten datarlas entre 1,8-1,2 Ma (Bienvenido Martínez-Navarro et al, 2015).
En Fuente Nueva 3 y Barranco León 5, aparecieron restos modo 1, datados en 1,4-1,2 Ma. En Fuente Nueva hay evidencias de acción humana sobre herbívoros para consumir carne y tuétano.
En Venta Micena:
VM-0, fragmento parietal-occipital, presumiblemente de un niño. Se trata de la parte posterior de un cráneo de un niño de no más de cinco años, datado en 1,3-1,2 Ma. Al ser un resto tan fragmentario y presentar una cresa prominente, ha generado mucha controversia. Para Martínez-Navarro (2002) corresponde a una hembra de bóvido o cérvido. En 2003 se halló en Francolí (Tarragona) el esqueleto de un niño, en una necrópolis del siglo IV, con la misma cresta (Campillo et al, 2006), aunque Martínez-Navarro (2008) afirma que las morfologías de la sutura no se corresponden. Según el análisis inmunológico efectuado por Concepción Borja et al (1997), el fragmento corresponde a un homínido.
VM1960, diáfasis humeral juvenil
VM3961, parte intermedia de un húmero adulto.
Barranco León. Ubicación, estratigrafía y dataciones. Agustí et al, 2015.
Francesc Ribot et al (2015) han estudiado la posición estratigráfica y espacial de BL5-0 y su morfología, llegando a la conclusión de que perteneció a un humano (parte mesial de un dm2 izquierdo), confirmando la opinión de Arribas y Palmqvist (2002).
Venta Micena. VM-0.
Hallazgos posteriores en Orce.
En Barranco León aparecieron fragmentos molares. Isidro Toro-Moyano et al, 2013, han descrito y datado el diente deciduo dm1 izquierdo de un homínido de unos diez años BL02-J54-100, Once boy,hallado en julio de 2002, asociado a 1.244 piezas líticas modo 1 y rectos óseos con fracturas y marcas de corte. La datación, de 1,4 Ma, convierte a este fósil en el resto homínido más antiguo de Europa Occidental. En las excavaciones de 2014 se descubrió un taller lítico.
Francesc Ribot et al (2015) consideran que BL5-0 y BL02-J54-100 posiblemente podrían ser contiguos, correspondiendo con la desarticulación de un humano fallecido.
BL02-J54-100. El resto homínido más antiguo de Europa Occidental. Jordi Mestre. IPHES.
María Patrocinio Espigares et al (2013) han investigado los restos de una carcasa de elefante hembra (Mammuthus meridionalis) en Fuente Nueva-3 cuya extracción finalizó en 2003. La carcasa apareció incompleta (le faltan las extremidades y el cráneo), rodeada de 17 lascas de sílex y 34 coprolitos de hiena gigante caricorta (Pachycrocuta brevirostris), lo que sugiere un comportamiento competitivo entre los homínidos y esta especie. Los coprolitos aparecieron en los espacios que hubieran correspondido a las extremidades y los autores interpretan que los homínidos llegaros los primeros al cadáver, cortaron y se llevaron las patas y el cráneo, marchándose lo más rápido que pudieron. Después las hienas acabaron con el resto del cadáver.
Yacimientos de Atapuerca
Sima del Elefante. Estratigrafía.
Sima del Elefante
En la cueva de la Sima del Elefante, Sierra de Atapuerca, España, se han hallado los siguientes fósiles humanos:
ATE 9-1, en 2007, un fragmento de mandíbula datado en 1,2-1,1 Ma.
La mandíbula no pudo ser atribuida a ninguna de las especies hasta entonces descritas. Incluye la región de la sínfisis, donde convergen las dos ramas mandibulares, la derecha conservada hasta el alveolo del primer molar y la izquierda hasta los alveolos del primer y segundo molar.
La parte de la mandíbula es primitiva.
La parte trasera es derivada.
El desgaste dental es muy severo. Para compensarlo, los dientes sobre erupcionaron y descalzaron.
Acumulaciones de cemento en las raíces dentales (hipercementosis).
Presencia de infecciones.
Dientes hacia fuera.
Utilización de palillos para higiene dental.
Del estudio del cálculo dental se deduce el consumo de plantas y el procesamiento de materias primas vegetales. Los restos de polen y fragmentos de insecto son consistentes con un entorno boscoso. No hay evidencias de cocinado de alimentos (Hardy et al, 2016).
El microdesgaste dental no oclusal presenta una alta densidad revelando el consumo de alimentos duros con desgaste por hueso triturado y arenilla, con un oportunismo alimenticio de amplio espectro que incluye carne y vegetales. Este patrón está presente también en los ejemplares de Homo antecessor (Pérez-Pérez et al, 2017).
Un premolar inferior de leche.
ATE 9-2, Una falange proximal del quinto dedo de la mano izquierda de un adulto. Robusta. (Carlos Lorenzo et al, 2014).
En 2011-2013 se encontraron restos líticos y fósiles animales con marcas de corte de aproximadamente 1,2 Ma y en 2013 un fragmento de cuchillo de sílex, que constituye la evidencia humana más antiguo en Europa Occidental, datado en 1,3 Ma. En 2014, se recuperó un lasca de sílex neógeno datada en 1,3 Ma. La tecnología corresponde con el Modo 1, muy pobre en cuanto diversidad y energía invertida en la producción de herramientas. Esta tecnología está relacionada con estrategias de subsistencia oportunistas que se centraron en el aprovechamiento de los cadáveres que habían caído en las cavidades (Andréu Ollé et al, 2013). La materia prima es de procedencia alóctona. Los restos de fauna sugieren abundantes zonas de agua, con hipopótamos y águilas pescadoras, prados abiertos con abundantes caballos, bóvidos, cérvidos y rinocerontes y condiciones templadas.
ATE 9-1
Hallazgo de la Sima del Elefante datado en 1,4 Ma, el resto más antiguo de Europa relacionado con homínidos
Otros yacimientos
La mayor parte, corresponden a un periodo de mejora climática marcada por el subcrón Jaramillo (1,07-0,99 Ma).
En Lezignan-la-Cébe (Hérault), Francia hallazgos líticos datados en 1,57 Ma.
En el nivel IV del Alto de las Picarazas (Andilla y Chelva, Valencia, España) con una datación de 1,5 Ma han aparecido herramientas líticas del Modo I en sílex y cuarcita, y además restos de fauna (caballos y bisontes) con marcas de corte. Huesos de conejo muestran, además de las marcas de corte con útiles líticos, marcas de dentición humana. De confirmarse la existencia de huesos quemados, estaríamos ante la evidencia de la producción del fuego más antigua de la Península Ibérica.
Kurgan Cimbal, Bogatiry y Rodniki, en Rusia. 1,2-0,8 Ma. Shchelinsky et al, 2003.
En el Valle Medio del Loira, Francia (Despriée et al, 2010):
El yacimiento lítico de Soleihac (Haute-Loire, Francia) está datado en 1,07-0,99 Ma.
En Grotte du Vallonnet, (Roquebrune-Cap Martin, Alpes Marítimos, Francia) hallazgos líticos datados en 1,05-1 Ma (de Lumley et al, 1988).
En los niveles TD3-TD4 de La Gran Dolina (Atapuerca) aparecieron algunas herramientas líticas (1 Ma). En 2013, se encontró un "chopper" de calcita tallado en uno de sus lados con el fin de cortar o descarnar animales.
En El Pino, Carrascosa del Campo, Cuenca, se ha comunicado el hallazgo de núcleos de cuarcita de cerca de 1 Ma.
Happisburgh, Norfolk, Inglaterra, es el primer asentamiento conocido al norte del paralelo 45. Hallazgos líticos y huellas humanas (Nick Ashton et al, 2014). Datado en 0,99-0,78 Ma, cuando la isla estaba unida al continente. Descubierto por Simon Parfitt et al (2010). Para Chris Stringer, los autores quizá pertenecieran al Homo antecessor.
Dursunlu, Turquía. 0,99-0,78 Ma. Gülec et al, 2009.
Korolevo, Ucrania. 0,99-0,96 ka. Koulakovska et al, 2010.
En Vallparadís (Terrasa, Barcelona) se han encontrado herramientas
modo 1 bipolares olduvaienses datadas en 0,9 Ma, con restos de fauna asociados. Algunos restos de herbívoros presentaban marcas de corte y fracturaciones antrópicas, demostrando así un acceso primario de los humanos a las carcasas de los animales en relación al resto de grandes carnívoros.
Primeros poblamientos humanos de Iberia. Pueden identificarse dos dispersiones faunísticas diferentes. Gibert et al, 2015.
En Cueva Negra, Caravaca de la Cruz, Murcia, apareció tecnología achelense datada en 0,9 Ma, con restos de fauna quemada y trozo de sílex deformado por el fuego. Son los restos de fuego controlado por humanos más antiguos de Europa.
Cueva Victoria, Cerro de San Ginés de la Jara, a 3 km del Mar Menor y a 13 km al este de Cartagena. Hallazgos datados entre 0,99-0,78 Ma. En el mismo lugar se hallaron restos del primate Theropithecus oswaldi. Esta dispersión africana de fauna, junto con la primera presencia del achelense en Cueva Negra apoyan una vía de entrada en Europa por Gibraltar (Gibert et al, 2015).
CV-0, falange.
CV-1 y CV-2, fragmentos de húmero.
En Ca'Belvedere di Monte Poggiolo (Forli, Emilia Romagna, Italia), hallazgos líticos datados en 0,85 Ma. Arzarello y Peretto, 2010.
Dorn-Dürkheim, Alemania, DD31 más de 0,8 Ma. Hallazgos líticos.
Pakefield, Inglaterra, artefactos de sílex datados en 0,7 Ma.
En Cúllar-Baza 1 (Granada, España), una escasa colección de piezas de sílex, y en menor medida de cuarcita, que se enmarcan dentro de la tecnología Modo I-Olduvayense datada entre 0,78-0,65 Ma.
En Solana del Zamborino, Granada, España, industria lítica en su mayor parte realizada con cuarzo y cuarcita aunque también se encuentran instrumentos de sílex, entre los que destaca un gran bifaz desconocido hasta el momento en Andalucía. En 2009 Gary Scott y Luis Gilbert realizaron un estudio geocronológico en el Berkeley Geochronology Center, publicado en la revista Nature, con el que descubrieron que el yacimiento era más antiguo de lo que se creía, situando su cronología de 0,76 Ma.
Sin datación precisa, en la Península Ibérica, disponemos además de los siguientes yacimientos:
El Aculadero (Puerto de Santa María, Cádiz).
El Rompido, Huelva.
Magoito, Mironço, Aldeia Nova y Açafora, en los alrededores de Lisboa.
Peteira (Tuy), Tomiño-Campo de Fútbol (Pontevedra), Chan de Vide – “Minas del Condado 3 y 5” (Pontevedra), Montes de Oleiros (Salvaterra de Miño y de As Neves), en las terrazas del tramo final del río Míño.
Monfarracinos y Toro, en las terrazas del Duero.
Hallazgos aislados en el entorno de Valladolid, en las terrazas del Pisueerga.
Perilla del Castro y Betrocino, en las terrazas del Esla.
Hallazgos en las terrazas del Tormes.
Pueblanueva, en las terrazas del Tajo.
Mas d’en Galí (Medinyà, Girona), Costa Roja (Sant Julià de Ramis, Girona) y el conjunto de Puig d’en Roca (Girona), en el curso medio del Ter.
Especiación de Homo antecessor. José María Bermúdez de Castro, 2013.
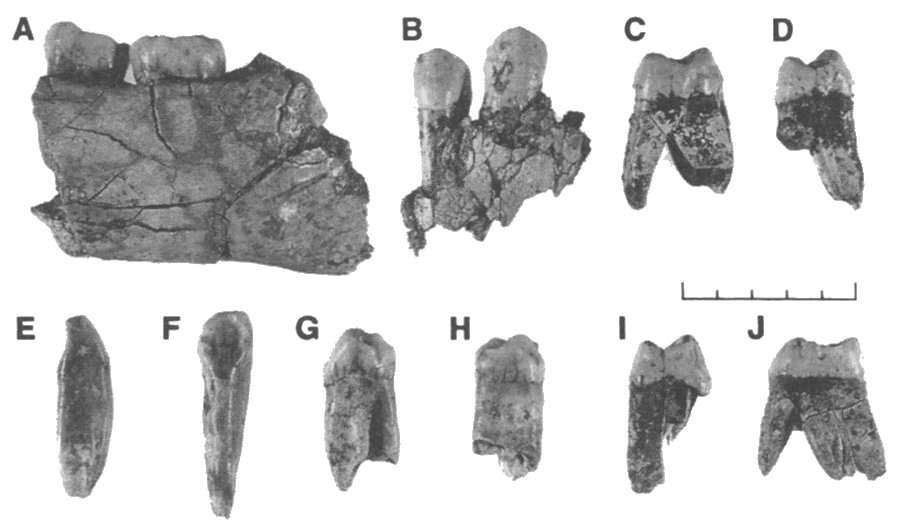
A partir de la realización de una cata de 7 m2 en 1976, se recogen de la Gran Dolina de Atapuerca (Burgos) restos que alcanzan más de 0,9 Ma (J.M. Parés et al, 2013). El nivel 6, del que proceden los hallazgos, alcanza los 2,5 metros de espesor y está formado por una colada con escasa matriz arcillosa. TD6-2, el estrato Aurora (nombre de la descubridora Aurora Martín Nájera), contenía ca 170 fósiles de Homo antecessor correspondientes a 10-15 individuos casi todos menores de quince años; la mayoría menores de dos años con algunos bebés.
Dataciones por ESR proporcionaron una edad entre hace 0,77-0,85 Ma para el límite superior de TD6 y de 0.91 ± 0.25 Ma para el límnite inferior de TD4. Análisis de lumniscencia, proporcionaron una edad de TD6 de hace 846 ± 57 ka. La combinación de datos obtenidos por ESR, luminiscencia, bioestratigrafía y resultados paleomagnéticos soportan una edad post-Jaramillo para la capa TD4 de Gran Dolina (Claudia Álvarez-Posada et al, 2018). La datación directa por ESR del diente ATD6-92 ha producido unas fechas entre hace 0,949-0,772 ka (Duval et al, 2018).
ATD6-96. Vista oclusal y lateral.
Sobresalen:
ATD6-5. Holotipo. Media mandíbula con sus tres muelas de un individuo joven. M3 está saliendo torcida, lo que debió originar dolor de muelas e incapacidad de morder. Sin los rasgos característicos de los preneandertales y pocos rasgos parecidos a los del ergaster.
ATD6-15. Restos parciales de la cara de un niño entre 10 y 11,5 años de edad. (Bermúdez de Castro et al 1997). Conserva el torus derecho y parte de la región de la glabela. Senos frontales muy desarrollados. La capacidad craneal pudo llegar a 1.000 cc.
ATD6-19. Pequeño fragmento cigomaxilar de un adulto, con un tubérculo cigomaxilar de las mismas características que el de ATD6-58, que se proyecta hacia fuera 2 mm (Arsuaga et al, 1999).
ATD6-22. Rótula derecha.
ATD6-38. Muy similar en tamaño y forma a ATD6-69. Su topografía sugiere claramente la presencia de una fosa canina (Arsuaga et al, 1999).
ATD6-43, fragmento de radio casi completo. El índice braquial y las proporciones son más próximos a los del ergaster y a los de los humanos modernos que a los de preneandertales y neandertales.
El ATD6-50 clavícula muy larga y relativamente delgada, con una curvatura del eje pronunciada y epífisis relativamente pequeñas, similar a la de los neandertales (Carretero et al, 1999).
ATD6-56. Rótula izquierda asociada a la rótula derecha ATD6-22. Presenta evidencia de un traumatismo.
ATD6-58. Fragmento cigomaxilar izquierdo de un adulto. Exhibe una notable fosa canina así como un gran tubérculo cigomaxilar situado en posición maxilar que se proyecta hacia fuera 3,3 mm (Arsuaga et al, 1999).
ATD6-69, El Chico de la Gran Dolina (indviduo 3). Gran parte del lado izquierdo de la cara, excepto las apófisis frontales de los huesos cigomáticos y maxilares. Conserva I2 derecho y P3, M1, M2 y M3 izquierdos permanentes. C y P4 derechos estaban finalizando el proceso de erupción. M2 izquierdo, todavía incluido en su cripta mientras que el germen de M3 se mantuvo unido al hueso por debajo de la cripta de M2. La edad dental, de acuerdo con los estándares modernos, se ha estimado en 10-11 años. Morfología totalmente moderna, incluyendo el patrón de desarrollo dental (Bermúdez de Castro et al, 1999), con rasgos más cercanos a los de los cráneos de Zhoukoudián y Dali y a los de Cromañón que a los neandertales (Rodrigo S. Lacruz et al, 2013, Sarah E. Freidline et al, 2013).
ATD6-92, fragmento de diente de un varón. Sobre este ejemplar se efectuó la primera datación por ESR de antecessor (Duval et al, 2018) y se obtuvo el proteoma humano más antiguo hasta la fecha (Welker et al, 2020).
ATD6-95 y ATD6-70+107 astrágalo izquierdo casi completo y segundo metatarsiano derecho asignados tentativamente al mismo homínido, un macho adulto. El astrágalo es largo y alto, con una tróclea larga y ancha y con un cuello proporcionalmente corto.
ATD6-96. Fragmento izquierdo de la mandíbula de una mujer muy joven. Exhibe un tubérculo pterigoideo hipertrofiado, una característica incluida en la lista de apomorfías neandertales (Carbonell et al, 2005; Rak et al, 1994; Weaber, 2009).
ATD6-121. Húmero infantil.
ATD6-124. 3/4 partes de un cuarto metatarsiano derecho. Según L. Martín-Francés et al (2013) el hueso estaba roto y se estaba soldando en el momento del fallecimiento del individuo. La presencia de esta fractura sugiere que la correspondiente población recorría grandes distancias o se desplazaba frecuentemente por un terreno desigual.
ATD6-148. Rafa. Húmero izquierdo de adulto muy joven vigoroso. Quebrado para obtener la médula. Marcas de descarnado y destrozo de los cóndilos para separar las masas musculares. Al igual que ATD6-121, exhibe una gran fosa del olécranon y unos pilares laterales y mediales muy finos, similares a los de los homínidos del Pleistoceno Medio europeo, los neandertales y Bodo (Carretero et al, 2009; Bermúdez de Castro et al, 2012).
ATD6-100/168. Un gran fragmento de parietal, el mayor recuperado hasta el momento del neurocráneo de antecessor. La morfología de las fracturas es compatible con un trauma peri-mortem. Las zonas parietales superiores son planas. Están presentes un gran foramen parietal y un foramen accesorio pequeño. Los vasos meníngeos medios no están particularmente desarrollados, pero se distribuyen en ambos sectores anterior y posterior, con una vascularización parietal procedente de la rama posterior. Los vasos meníngeos muestran múltiples conexiones de menor importancia con los sistemas pericraneal y diploico. El diploe no está particularmente desarrollado, y no se detectan grandes canales diploicos. El hueso es delgado. Esta descripión sugiere que el hueso perteneció a un juvenil. Bruner et al, 2016.
En 2013 se acabó de liberar una escápula infantil hallada en 2005, correspondiente a un infante entre 4-6 años.
También, más de 800 artefactos líticos:
En el nivel TD4, un chopper que fue utilizado para descuartizar huesos de animales, datado en 1 Ma. El techo del nivel TD4 había deparado ya varias herramientas de cuarcita manufacturadas a partir de cantos del río Arlanzón.
En TD5, tres artefactos, entre ellos un núcleo de cuarcita y una lasca de sílex.
En TD6, asociada a los restos humanos, aparece una abundante tecnología lítica (n>300) con núcleos y lascas (industrias arcaicas o Modo I); la materia prima utilizada es el sílex, cuarzo, cuarcita, caliza y arenisca.
Pese a que la tecnología no varió sustancialmente existieron dos fases principales de ocupación humana: En una primera, los Homo utilizaron muy marginalmente la cavidad pero poco a poco su presencia se va haciendo más intensa hasta que en las fases medias y finales se asienta un campamento base, en el que se incrementan notablemente las actividades, incluyendo los eventos de canibalismo (Marina Mosquera et al, 2018).
Los cráneos tienen más de 1.000 cc y el torus supraorbital de doble arco es claramente distinguible del erectus. Menor grado de encefalización que en sapiens. Cerebro lateralizado. Los dientes son muy parecidos a los del grado erectus. Está presente la fosa canina, ausente en los erectus de Asia. El M1 inferior es más similar al del HAM que al de la población de la Sima de los Huesos y los neandertales (Martínez de Pinillos et al, 2015). Un estudio de los dientes deciduos ha revelado características en mosaico con algunos rasgos avanzados que anticipan la morfología derivada de los dientes europeos del Pleistoceno (José María Bermúdez de Castro et al, 2017). Los molares permanentes exhiben un esmalte grueso, tanto en la corona como en el lateral. Esta condición se comparte con la mayoría de las muestras de Homo extintos y existentes, a excepción de los neandertales y algunos especímenes aislados. Sin embargo, mientras que el porcentaje de dentina en la corona total se asemeja a los valores modernos bajos, el porcentaje en el lateral tiende a ser mucho más alto, más cercano al patrón neandertal. Los mapas de distribución del esmalte molar revelan un patrón de distribución relativa que es más similar a la condición neandertal (con el esmalte más grueso en la periferia de la cuenca oclusal) en lugar de otros especímenes fósiles y humanos modernos (con esmalte cuspal más grueso) (Martín-Francés et al, 2018). Según García-Campos et al (2018), los dientes exhiben grandes dimensiones de dentina coronal y radicular, así como un patrón esmaltado delgado, que tradicionalmente se ha considerado una autapomorfia neandertal. Para Martinón-Torres et al (2019), la evaluación de la muestra dental respalda la validez taxonómica de antecessor.
En cuanto al esqueleto postcraneal, H. antecessor se parece más a los HAM que a los homínidos europeos de mediados y finales del Pleistoceno (Carretero et al, 1999; Lorenzo et al, 1999), aunque algunos rasgos también se comparten con estos últimos (Pablos et al, 2012). Los metatarsianos no difieren significativamente de los correspondientes a los humanos modernos, neandertales y ejemplares de la Sima de los Huesos.
En resumen, H. antecessor se caracteriza por una combinación única de características plesiomórficas y derivadas (Bermúdez de Castro et al, 2017).
Plesiomórficas:
La dentición: desarrollo de un cíngulo en premolares y molares, crenulaciones del esmalte profusas, asimetría de la corona del primer premolar inferior, gran tamaño en molares y sistemas radiculares complejos y robustos. Ausencia de espacio retromolar. M3 parcialmente cubierto por la rama mandibular. Fuerte tubérculo molar en el dm1 deciduo. Presencia de cúspides mesial y lingual en el canino superior deciduo. Las caras de los premolares mandibulares muestran crestas y surcos marginales mesiales y distales, que se conectan con el cíngulo. La forma de la corona del P3 mandibular es fuertemente asimétrica, con un notable talónido y un gran polígono oclusal. El P4 mandibular muestra un contorno asimétrico, con un metacónido desplazado mesialmente con talónido abultado y un amplio polígono oclusal. P3 y P4 mandibulares muestran un sistema radicular complejo, con una raíz mesiobucal con dos canales de pulpa y una raíz distolingual con un solo canal. M1<M2 para las series molares superior e inferior. M1 y M2 mandibulares exhiben un patrón Y de los surcos bucales y linguales que separan las cinco cúspides principales. El esmalte de la superficie oclusal de los dientes poscaninos está moderada o notablemente crenulado. Cuello radial largo tanto desde un punto de vista absoluto como relativo.
Los rasgos endocraneales de los parietales.
Frente huidiza y toro supraorbital de doble arco.
El proceso estiloides esta fusionado al basicráneo.
Suelo nasal inclinado.
Ausencia de mentón.
Posición del foramen mental al nivel de P3-P4 mandibular.
Baja posición de la línea milohioidea en relación al margen alveolar en el nivel M3 mandibular. Trayectoria paralela de la línea milohioidea en relación al margen alveolar.
Fosa pterigoidea poco profunda.
Apomorfias compartidas con Hominini posteriores.
Torus supraorbital doblemente arqueado.
Los parietales muestran un aplanamiento parasagital de las superficies superiores.
Posición de la prominencia lateral al nivel del talónido del M2 mandibular. Ausencia de prominencia alveolar. Trigonum postmolare inclinado oblicuamente. Fosa masetérica poco profunda. Fosa subalveolar moderadamente profunda. Perfil regular del gonion. El ratio longitud/anchura de la arcada alveolar mandibular está claramente por debajo de cien.
Incisivos permanentes mandibulares bucolingualmente expandidos con respecto a H. habilis s.s. y otros Hominini africanos y euroasiáticos del Pleistoceno Temprano. P3>P4 para el área coronal de los premolares. Dientes poscaninos más pequeños que los de H. habilis s.s. y en el rango de H. ergaster, H. erectus s.l., y Hominini europeos del Pleistoceno Medio. M3 mandibular notablemente reducido con respecto a M1 y M2.
Características derivadas, compartidas con los neandertales y los HAM.
Borde superior convexo de la escama del temporal, y una posición anterior del conducto incisivo, que es casi vertical (Arsuaga et al, 1999).
Planum alveolare ausente o débil.
Capacidad craneal, estimada en más de 1.000 cc.
Características derivadas, compartidas con los HAM.
La cara. Más moderna que la de los homínidos africanos del Pleistoceno Inferior, caracterizados por un prognatismo pronunciado. Es plana, con un nasal saliente, maxilar poco proyectado y mandíbula delgada. Fosa canina, consecuencia de una reorientación hacia delante y abajo de las placas infraorbitarias cuyo borde inferior parte de una posición horizontal y alta.
Posición medial del punto más profundo de la hendidura mandibular.
Rótula relativamente estreha, con indices rotulianos altos.
Características derivadas compartidas con los neandertales y la población de la Sima de los Huesos.
Presencia de un tubérculo pterigoideo medial.
Polígono oclusal reforzado y comprimido y un contorno externo sesgado de la M1 permanente con protuberancia abultada del hipocono.
La región mastoidea es una reminiscencia de la de los neandertales.
Incisivos superiores permanentes muestran forma de pala, especialmente I2.
Presencia de cresta media del trigónido.
Fosa olécranona grande y pilares medianos y laterales muy finos
Clavícula muy larga en términos absolutos, relativamente delgada (bajo índice de robustez), con pronunciada curvatura del eje y epífisis relativamente pequeña.
Tróclea del astrágalo relativamente estrecha.
MORFOLOGÍA FACIAL
ESCAMA TEMPORAL
Homo habilis
Homo rudolfensis
Cara plana o cóncava.
Placas infraorbitales dispuestas coronalmente, verticales u orientadas hacia arriba.
La apertura nasal no se sitúa por delante de las placas infraorbitarias.
Borde bajo y rectilíneo.
Grado erectus
Cavidad nasal ligeramente más adelantada respecto a las placas infraorbitarias. Prognato.
Huesos nasales dispuestos más horizontalmente.
Zona de crecimiento óseo bajo la nariz. Osteoblastos. Topografía subnasal plana.
Borde bajo y rectilíneo.
Homo antecessor
Placas infraorbitales orientadas coronalmente e inclinadas hacia abajo, con fosa canina.
Resorción ósea bajo la nariz. Osteoclastos. Topografía subnasal convexa. Tubérculo pterigoideo medial hipertrofiado.
Borde elevado y arqueado.
Homo neanderthalensis
Cara de aspecto apuntado.
Cavidad nasal situada más hacia delante que en las demás especies.
Huesos nasales dispuestos horizontalmente.
A ambos lados de la nariz, las paredes de la cavidad nasal y las placas infraorbitarias forman una única superificie sin fosa canina, orientadas oblicuamente. Tubérculo pterigoideo medial hipertrofiado.
Borde elevado y arqueado.
Homo sapiens
Con fosa canina.
Resorción ósea bajo la nariz. Osteoclastos.
Borde elevado y arqueado.
Homo antecessor. ATD6-69 y ATD6-15.
Estatura, proporciones y peso
Una gran mayoría de individuos medirían entre 160 y 180 cm, siendo el promedio de los machos algo superior al promedio de las hembras, como sucede en la actualidad en nuestra especie. En función de la tibia, el fósil original que se exhibe en el MEH, mediría unos 175 cm. Proporciones corporales: similares a las de Homo sapiens, pero con el tronco más ancho. Peso corporal: un 30% mayor que sapiens, debido a una mayor masa esquelética y muscular y a una mayor anchura del tronco.
Las diferencias de peso y estatura entre machos y hembras serían similares a las del sapiens.
Ciclo de vida
Parto similar al de Homo sapiens en cuanto a la fisiología, movimientos del niño y modo de presentación, pero con un tránsito más holgado por el canal del parto, debido a las mayores dimensiones de la pelvis. Lactancia: entre tres y cuatro años. Desarrollo probablemente casi tan largo y complejo como en nuestra especie, con infancia, niñez, fase juvenil y adolescencia, incluido un estirón puberal de intensidad similar a la nuestra.
Longevidad: menor que la de Homo sapiens. Su estilo de vida no permitiría una vida más allá de los 40 o 45 años. La esperanza de vida al nacimiento sería de entre quince y veinte años.
Posibles causas de muerte: parto, infecciones de heridas y de la cavidad bucal, traumatismos producidos por golpes accidentales o por violencia intra-específica, predadores.
Modo de vida y dieta
Se trata de una asentamiento de larga duración (un campamento base), pero la antropofagia sugiere una cierta sincronicidad en la deposición, de unas decenas o cientos de años. Campaña et al (2016) proponen un origen alóctono: la actividad de los Homo se habría desarrollado fuera de la cueva, pero muy cerca de ella. la excelente conservación tafonómica de los huesos indica un tiempo muy corto a la exposición externa, un transporte rápido y en distancias muy cortas. Una consecuencia de esta hipótesis es que la datación de los correspondientes fósiles podría ser más antigua de la que se supone actualmente. Cazadores y recolectores de pequeños animales, huevos y todo tipo de plantas silvestres comestibles. Estos seres utilizaban distintas estrategias para cazar, se organizaban en clanes de entre 15 y 40 individuos jerarquizados y transportaban las piezas de diferentes formas, seleccionándolas o no según su tamaño, la hora del día, la distancia a la base o la presencia de otros carnívoros en el entorno. De estos hechos podemos inferir una gran complejidad social con reparto de tareas (Palmira Saladié, 2011). Contactos esporádicos con otros grupos para evitar la endogamia. La presencia de costillas (que son consumidas por los grandes depredadores) y de marcas de corte realizadas con cuchillos de piedra permite deducir que mataban ellos mismos a estos animales. Los restos humanos presentan las mismas huellas de tratamiento con herramientas de piedra que los restos de animales de lo que se deduce canibalismo (Palmira Saladié et al, 2015). Según Palmira Saladié et al (2012) los homínidos de TD6 realizaban ataques de bajo riesgo a los miembros de otros grupos para defender el acceso a los recursos dentro de sus propios territorios y tratar de ampliar sus territorios a costa de los grupos vecinos. Según Guillermo Rodríguez-Gómez et al (2013), el entorno era muy rico en recursos, con un nicho vacío para los predadores, y los homínidos podían aprovisionarse de forma continua de abundante carne y grasa por lo que hay que descartar el hambre como causa del canibalismo. Podría tratarse de víctimas de enfrentamientos territoriales de carácter estructural. Rosa Huguet el al (2013) han estudiado los restos de fauna recuperados en La Gran Dolina (niveles 3-4 y 6) y en la Sima del Elefante (niveles 9-14). Abundan los restos de fauna como producto de la práctica cinegética, con abundantes marcas de corte en los restos esqueléticos, que han sido fragmentados sistemáticamente por la acción antrópica. Las estrategias de subsistencia tuvieron un éxito suficiente como para permitir que los grupos de homínidos sobreviviesen y se reprodujesen en número suficiente. Por lo tanto, estos primeros seres humanos tendrían la capacidad de mantener una ocupación continua de Europa.
Otras conclusiones:
Se constata un acceso primario de los homínidos (anterior al de los carroñeros) y no se ha constatado acceso secundario (Palmira Saladié et al, 2014)
Se efectuaban todas las tareas de carnicero.
No se seleccionaban los animales.
Los homínidos desarrollaron un comportamiento complejo de estrategias de subsistencia oportunistas y generalistas con un vasto conocimiento y aprovechamiento del territorio.
De su higiene sabemos que usaban palillos de dientes para la limpieza de los espacios interdentales. Usaban abrigos de piel para evitar el frío.
Campamentos con cabañas de madera y quizás pieles de animales, pero también uso de abrigos y vestíbulos de cuevas.
La industria lítica hallada es pre-achelense, Modo
1 o olduvaiense evolucionado, con ausencia de hachas de mano. El conjunto lítico es rico y diversificado. Los bloques de sílex para núcleos fueron amontonados de forma intencional. Se observa la representación completa de la cadena operativa que exige la fabricación de los instrumentos líticos, es decir, la existencia de núcleos, lascas, útiles, así como de fragmentos producto de la actividad de talla. La
actividad productiva de taller en el sitio se ha confirmado mediante diversos
remontajes. Predominan las bases positivas, de las que son minoría las
retocadas. Se ha comprobado su utilización para cortar carne, preparar huesos
para romperlos, descortezar y raer madera y posiblemente se utilizasen también
para tratar la piel. Se han documentado actividades de caza y los primeros rastros de canibalismo en la prehistoria (Andréu Ollé et al, 2013)
Del conjunto faunístico asociado se deduce un
ambiente de bosque abierto o con pradera en proximidad y un remanso o laguna
muy cercano, con un clima cálido, de humedad estacional; unas condiciones
climáticas similares a las de los actuales inviernos en la zona. En este
tiempo, la cavidad de la Gran Dolina fue un lugar de consumo.
Los restos de polen sugieren una recuperación de
temperaturas después de una fase fría.
La clasificación de los restos de La Gran Dolina.
Lo habitual para los restos europeos de transición era
clasificarlos como heidelbergensis.
Para Carbonell (1995), se trataría de un taxón diferente al erectus (que se extingue) y al ergaster (que daría lugar al sapiens).
Howell le asignó el paleodemo Atapuerca-Gran Dolina.
El equipo de Atapuerca propuso la nueva especie Homo antecessor (Carbonell et al, 1997), con una topografía mediofacial completamente moderna y otros
rasgos que lo apartaban de erectus,
ergaster y neanderthalensis.
Está relacionada con los pobladores del
Pleistoceno antiguo de China. Ambos demos procederían del grupo que abandonó
África hace 1,9 Ma. Para comprobar la relación sería necesario hallar fósiles
de este periodo en el este de Europa y oeste de Asia.
María Martinón-Torres et al (2007) a partir de un estudio fenétido y cladístico de la dentición diferencian entre:
Un bloque africano (A. africanus, A. afarensis, H. habilis, H. ergaster y H. mauritanicus).
En la dentición posterior, aumento significativo de cúspides accesorias y rasgos que complican profusamente la superficie oclusal.
Un bloque euroasiático (H. antecessor, H. erectus s.s., H. heidelbergensis y H. neanderthalensis).
En la dentición anterior, forma en pala triangular, con una convexidad labial pronunciada y una superficie oclusal en "V" por el desarrollo de un surco estrecho secundario.
En la posterior, rasgos derivados de la reducción dental como la pérdida de cúspides en molares, simplificación de las superficies oclusales y aparición de patrones de surco diferentes del driopitecino. Cresta media del trigónido continua con una fovea anterior corta y profunda. Cresta transversa continua en segundos premolares inferiores.
H. sapiens y H. georgicus muestran una posición intermedia.
Esto sugiere que los senderos evolutivos de los continentes africano y euroasiático fueron independientes a lo largo del Pleistoceno Inferior y Medio. Ello implica un origen euroasiático para H. antecessor, quizá en el Levante Mediterráneo.
Para Emiliano Aguirre (2008) no
tendría relación filogenética directa ni con erectus ni con heidelbergensis
o sapiens. Aguirre (2000) es defensor de la teoría mutirregional de
evolución hacia el sapiens y cree que
rasgos faciales compartidos por antecessor,
erectus y sapiens indican un
aportegenético muy variado como
origen de los humanos actuales.
El rostro de antecessor combina características compartidas con el HAM con otras similares a las de los neandertales y grado heidelbergensis. Sarah E. Freidline et al (2013) han sugerido que algunos de los rasgos faciales que caracterizan a los últimos humanos modernos pueden haberse desarrollado varias veces en la evolución humana.
El equipo de Atapuerca considera a Homo atecessor un buen candidato a la ascendencia común de HAM y neandertales. Mendez et al (2016) han estimado que la separación entre neandertales y HAM se produjo hace 806-447 ka para el ADN-Y. Esta fecha cae dentro del rango de antecessor y fuera de heidelbergensis. Gómez-Robles et al (2013) señalan que ninguna especie conocida muestra una morfología dental compatible con la que se podría esperar para un antecesor común de neandertales y HAM.
José María Bermúdez de Castro y María Martinón-Torres examinan la hipótesis de la convergencia deSarah E. Freidline et al (2013) a la luz de los fósiles humanos recuperados en TD6. Destacan que basan su interpretación en la ausencia de una cara totalmente moderna en el Pleistoceno Medio africano, pero debido a la ausencia de fósiles, solo es posible tener en cuenta Kabwe y Bodo. Para este último, algunas características del infraorbitario permitirían alinearlo con H. sapiens. La falta de una muestra representativa, llama a la prudencia.
Como una alternativa a las interpretaciones de Sarah E. Freidline et al (2013), los autores proponen una cladogénesis del género Homo durante el Pleistoceno Temprano, que dio lugar a un aumento del tamaño craneal y a una morfología facial moderna.
KNM-WT 15000 representa la condición primitiva del clado Homo
H. antecessor deriva probablemente de este cladogénesis y representa una rama lateral confinada a Europa Occidental (Bermúdez de Castro et al, 2003; Carbonell et al, 2005; Martinón-Torres et al, 2007; Bermúdez de Castro y otros, 2012). H. antecessor comparte características derivadas con neanderthalensis, la población de la Sima de los Huesos, los Homo del Pleistoceno Medio de Chinay sapiens y estaría muy cerca de su ancestro común más reciente. Precisamente debido a esta posición filogenética, niegan una posible relación ancestro-descendiente con la especie ñ (José María Bermúdez de Castro et al, 2013; José María Bermúdez de Castro et al, 2014;José María Bermúdez de Castro et al, 2015).
El prognatismo mediofacial que puede observarse en los neandertales clásicos (Rak, 1986), y que también está presente en los homínidos del Pleistoceno de Europa Oriental (Arsuaga et al, 1999), representaría otra condición derivada que podría haber surgido a partir de una cara similar a la moderna (Arsuaga et al, 1999). Estos rasgos fueron retenidos por los neandertales (sinapomorfías) (Gómez-Robles et al, 2007; Bermúdez de Castro y Martinón-Torres 2012; Martinón-Torres et al, 2013).
El espécimen de Bodo presenta algunas sinapomorfias con neandertales y HAM en diversos grados y podría formar parte de uno de los linajes de la cladogénesis que en África dio lugar a H. sapiens y en Europa a H. neanderthalensis.
Welker et al (2020), tras analizar el proteoma del esmalte dental de Homo antecessor (ADT6-92) y Homo erectus de Dmanisi (D4163), han obtenido evidencia de que antecessor es un linaje hermano que se separó del ancestro común de humanos modernos, neandertales y denisovanos. Esta ubicación implica que la cara moderna de antecessor, similar a la de los humanos modernos, puede tener una ascendencia considerablemente profunda en el género Homo, y que la morfología craneal de los neandertales representa una forma derivada.
Cueva Negra del Estrecho del Río Quípar (España) 0,78 Ma, según datos bioestratigráficos (M. J. Walker et al, 2012). Fragmentos de de húmero y cúbito humano y dientes.
En España hay yacimientos líticos datados en más de 0,75 Ma en Barranco León, Fuentenueva-3, Monfarracinos, Galisteo, Talavera, El Espinar, Atapuerca y Huéscar.