Australopithecus afarensis, Johanson, White y Coppens, 1978.
- Holotipo: LH4. Mandíbula con dentición de un adulto.
- Yacimientos:
- Tanzania
- Laetoli
- Kenia
- Koobi Fora
- West Turkana
- Kantis (Mbua et al, 2016)
- Etiopia
- Dikika
- Omo
- Maka
- Hadar
- Middle Awash
- Woranso-Mille, Afar.
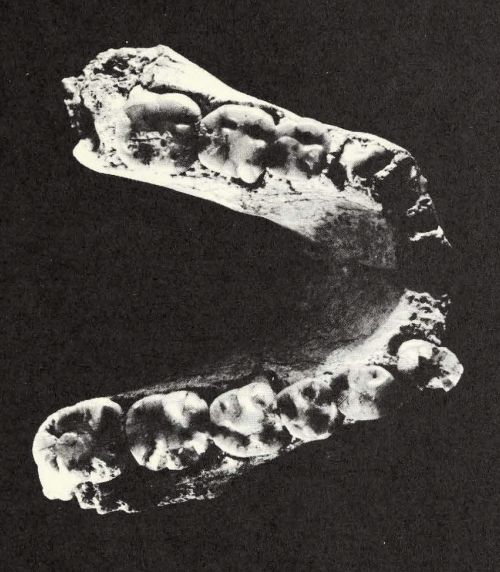 |
| LH 4, vista oclusal. (Johanson, et al. 1978). |
Los primeros restos descubiertos, a principios de la década de 1970, se identificaron con Homo y solo tras posteriores estudios se reconoció la nueva especie. Afarensis, deriva de Afar, la región de Etiopía de los primeros hallazgos. Las descripciones iniciales destacaron la similitud a los grandes simios cuello arriba y a los humanos cuello abajo, así como un fuerte dimorfismo sexual. En 1974 se descubrió AL-288-1, un esqueleto bastante completo que recibió el nombre de Lucy. Su fama empañó el descubrimiento al año siguiente de la primera familia, los fósiles de un grupo de individuos que murieron a la vez, en la localidad AL333. En 2006-2007, se describió el hallazgo de Dikika los fósiles poscraneales de un individuo joven que revelaron detalles desconocidos hasta el momento. En 2013, Donald Johanson anunció el hallazgo en Etiopía de un cráneo completo que corresponde a una hembra.
A. afarensis se distingue de A. anamensis por:
Cabe preguntarse en qué medida cabe incluir en la misma especie ejemplares de Laetoli y Hadar, muy separados entre sí, además de una distancia en el espacio de 1.800 km., y en el tiempo de 0,8 Ma.
- P3 con una corona más molarizada y simétrica con una segunda cúspide más frecuente (metacónido) y una orientación transversal con respecto a la hilera de dientes.
- Canino superior asimétrico con el hombro mesial de la corona más apical.
- dm1 molarizado, con talónido expandido bucolingualmente.
- Perfil del corpus anterior recto, más comúnmente vertical, con un segmento basal lleno y poca o ninguna inflexión inferomedial del corpus inferior bajo los caninos y premolares.
- Caninos inferiores posicionados medialmente al eje de la fila de poscaninos.
- Paladar equivalente o más ancho.
- Apertura nasal definida por márgenes agudos laterales y márgenes inferiores diferenciados, con el jugum canino con entidad propia en la topografía circumnasal.
- Meato auditivo externo más grande.
 |
| Ubicación de Laetoli |
Hallazgos
- Laetoli, Tanzania. 3,7-3,5 Ma. Dientes.
- Huellas de Laetoli
- Sitio G. Paul Abell ,1978. 3,7 Ma. Huellas dejadas por varios individuos bípedos:
- G1, individuo infantil.
- G2, individuo adulto.
- G3, superpuestas a G2.
- Sitio S. Masao et al, 2016. A unos 150 m del sitio G.
- S1 y S2, que se mueven en la misma paleosuperficie y en la misma dirección que los tres individuos documentados en el sitio G. Las huellas de G y S se consideran contemporáneas.
- La huella de S1 ("Chewie") mide 26 cm y pudo pertenecer a un macho de 165 cm y 45-48 kg. Las otras huellas, incluyendo las del sitio G, corresponderían a hembras y juveniles.
- Un macho caminando con varias hembras sugiere una estructura social cercana a la de los gorilas.
- Garusi I. Mandíbula.
- LH-2. Mandíbula.
- LH-4. Holotipo.
- LH-5. Mandíbula.
- LH-21. Esqueleto juvenil parcial fragmentario.
 |
| Estratigrafía de Laetoli |
 |
| KSD-VP-1/1. Australopithecus afarensis. |
- Woranso-Mille (WORMIL), Afar. Etiopía. Estudiado por Haile-Selassie.
- KSD-VP-1/1 Kadanuumuu. Muchos fragmentos postcraneales. Haile-Selassie et al, 2010. Yohannes Haile-Selassie y Denise F. Su (2015), han coordinado la publicación de los estudios sobre KSD-VP-1/1, Kadanuumuu, correspondiente a un macho adulto, hallado en Woranso-Mille, Etiopía, ca 40 km al norte de Hadar, Gona y Dikika, y datado en hace 3,6 Ma (Saylor et al, 2015).
- Kadanuumuu muestra la única tibia completa conservada de afarensis y apoya la hipótesis de que sus piernas eran relativamente largas, como en los humanos.
- Su tórax revela una postura completamente erguida (Latimer et al, 2015).
- Se han recuperado seis vértebras cervicales parciales (C2-C7), las más antiguas conocidas de los Hominini. Son más derivadas que las de Lucy y Selam y evidencian un régimen de carga ortrógado y un cuello muy móvil (Meyer, 2015).
- La escápula y la clavícula apoyan la interpretación de que el hombro de afarensis estaba más cranealmente orientado que el de los humanos, pero por otra parte el tamaño grande de la fosa infraespinosa y la musculatura de la clavícula y la escápula recuerdan a las de los humanos, sugiriendo que la extremidad superior empezaba a enfocarse en la manipulación. (Melillo, 2015).
- La pelvis es de aspecto moderno y difiere poco de la de los HAM (Lovejoy et al, 2015)
- Probablemente el hábitat estaba constituido por bosques densos con algunas áreas distantes de pastizales abiertos o matorrales (Su, 2015).
 |
| KSD-VP-1/1. Australopithecus afarensis. Reconstrucción de la pelvis. |
- Kantis, cerca de Nairobi, Kenya. 3,5-3,3 Ma.
- Algunos dientes probablemente de dos menores y un adulto y un cubito izquierdo (KNM-RK 53525) de un macho adulto. La fauna y los isótopos de carbono sugieren un entorno más abierto y con plantas C4 que el reportado para otros yacimientos de afarensis, con prados, una fuente de agua permanente y algunos árboles. Estos hallazgos amplían el rango de distribución de afarensis a las tierras altas al este del Valle del Rift (Mbua et al, 2016).
- Maka, Middle Awash, Etiopía. 3,4 Ma. Algunas mandíbulas, una de ellas casi completa.
- Fémur proximal.
- Húmero.
 |
| Ubicación de Hadar y Dikika |
- Hadar, Etiopía. 3,4-2,96 Ma. Miembros Sidi Hakoma, Denen Dora y Kada. 367 especímenes (90% del total de la especie). 57 mandíbulas de adulto o subadulto. 12 calvarias.
- AL 129-1 (Donald Johanson, 1973). 3,4 Ma. Porciones de ambas piernas, incluyendo una rodilla derecha muy pequeña, pero que aparentemente, pertenece a un adulto.
- AL 288-1, Lucy (Donald Johanson y Tom Gray, 1974). 3,2 Ma. Lucy fué una hembra adulta de 25 años. Se encontró el 40% de su esqueleto. La pelvis, el fémur y la tibia demuestran que era bípeda. Medía 1'07 cm de altura y pesaba 28 kg.
- AL 333 Site, The First Family (Donald Johanson, 1975). 3,2 Ma. Restos pertenecientes a 13 individuos de todas las edades. La talla de los individuos varía considerablemente, lo que ha dado lugar a interpretar la presencia de una sola especie o de varias.
- AL 333-1. Parte derecha mandibular de un infante.
- AL 333-105. Cráneo juvenil parcial.
- AL 333-160, un cuarto metatarsiano del pie izquierdo. Ward, Kimbel y Johanson (2011).
- Abundantes restos de miembros superiores, miembros inferiores y material axial.
- AL 400-1a, mandíbula inferior halladan entre 1974-1977 por Don Johanson. Está fechada en ca 3 Ma. Relativamente larga y estrecha, con molares traseros de tamaño moderado con patrón 'Y-5' e incisivos relativamente amplios.
- AL 417-1. Cráneo parcial.
- AL 438-1. Esqueleto parcial con asociaciones craneodentales.
- AL 444-2. (Bill Kimbel y Yoel Rak, 1991). 3 Ma. Hallado por Tim White. Se trata de un cráneo con el 70% de los huesos, correspondiente a un macho adulto grande, con capacidad craneal de 550 cc. De la comparación con Lucy se deduce un gran dimorfismo sexual.
- AL 822-1. Cráneo casi completo. Hallado en 2000. Probablemente de hembra por su pequeño tamaño.
- AL 1030-1. (Halszka Glowacka, William H. Kimbel y Donald C. Johanson, 2017). Fragmento del corpus mandibular izquierdo de un infante con dp3 y dp4 erupcionados. Son visibles las coronas de I1 e I2 no erupcionadas. Conserva la rañiz del ramus ascendente. Recuperado en 2002. Datado en hace ca 3,2 Ma.
- AL 1920-1. (Halszka Glowacka, William H. Kimbel y Donald C. Johanson, 2017). Mandíbula paracial de un juvenil que conserva completo el corpus derecho con el nacimiento del ramus ascendente y la región sinfisial con el corpus anterior izquierdo adyacente. Recuperado en 2012 con una datación de ca 3,35 Ma.
 |
| Estratigrafía de Hadar. Donald Johanson, 2016. |
 |
| AL 129-1. Australopithecus afarensis. |
 |
| AL 200-1 Australopithecus afarensis |
 |
| AL 444-2. Australopithecus afarensis |
- Koobi Fora, East Turkana, Kenya (Miembro Tulu Bor). 3,35 Ma.
- KNM-ER 2602. Calvaria parcial.
- Dikika (al sur y cerca de Hadar), Etiopía. 3,3 Ma.
- DIK-1/1, Selam, Bebé Dikika (Zeresenay Alemseged, 2000). Cráneo y resto de esqueleto bastante completo de una niña de tres años, según estimaciones a partir de la dentición. Hioides primitivo, con dificultad para emitir sonidos articulados debido a la presencia de sacos de aire. Bipedia incipiente y conservación de la trepa (DeSilva et al, 2018). Escápula similar a la del gorila. Muestra siete vértebras cervicales y sólo 12 vértebras torácicas como los seres humanos, en lugar de 13 como los simios africanos (Ward et al, 2017). Canales semicirculares parecidos a los de los simios africanos. Su volumen craneal corresponde a un 65-88% del típico de la especie. En los chimpancés a los tres años se alcanza el 90%, de lo que se deduce que, en nuestra línea evolutiva, la infancia estaba ya en proceso de prolongación.
- En la localidad DIK-55 fueron hallados dos huesos con marcas de corte, datados en 3,39 Ma (McPherron et al 2010 y Thompson et al 2015, pero Manuel Domínguez-Rodrigo, Travis Rayne Pickering y Henry T. Bunn 2011).
- Yonatan Sahle, Sireen El Zaatari y Tim D. White (2017) advierten de la posibilidad de que las marcas de corte atribuidas a los homininos arcaicos puedieran ser causadas por cocodrilos.
 |
| DIK 1/1 |
 |
| DIK 1/1 |
- Nefuraytu, Woranso-Mille, Afar, Etiopía, con una datación de hace 3,330-3,207 Ma. Haile-Selassie et al, 2016.
- NFR-VP-1/29, es una de las mandíbulas más completas asignadas a la especie hasta el momento. Es una de las mandíbulas de mayor tamaño atribuidas a Au. Afarensis por lo que probablemente perteneció a un macho. Presenta casi todas las características arcaicas distintivas documentadas para Au. Afarensis. Conserva los dientes izquierdos I2, P4 y M3; y los derechos P4 y M3.
- NFR-VP-1/2, es una corona completa sin erupcionar de un M1 inferior derecho.
- NFR-VP-1/58, es una corona de M2 o M3 inferior derecho.
- NFR-VP-1/96: Mitad lingual extremadamente desgastada de un canino inferior.
- NFR-VP-1/213: Es una corona de un M3 superior derecho.
- NFR-VP-1/214: pequeño fragmento de un hueso frontal.
 |
| Fósiles de Australopithecus afarensis procedentes de Nefuraytu, Woranso-Mille, Afar, Etiopía, con una datación de hace 3,330-3,207 Ma. a, b, c y d: NFR-VP-1/29. e: NFR-VP-1/2 f: NFR-VP-1/58 g: NFR-VP-1/213 |
HALLAZGOS CRANEALES
|
|||
Cráneos completos
|
Cráneos fragmentarios
|
Mandíbulas
|
|
HADAR
Etiopía
3,4-2,9
mda
|
AL
333-105 (juvenil)
AL
444-2
|
AL
58-22
AL
162-28
AL
199-1
AL
200-1
AL
288-1
AL
333-1
AL
333-2
AL
333-45
AL
333-86 (juvenil)
AL
333-125
AL
417-1
AL 438
|
AL
128-23
AL
145-35
AL
188-1
AL
198-1
AL
207-13
AL
266-1
AL
277-1
AL
288-1
AL
311-1
AL
333w-1
AL
333w-12
AL
333w-60
AL
400-1
NFR-VP-1/29
|
LAETOLI
Tanzania
3,6 mda
|
LH 21
|
LH 4
|
|
KOOBI
FORA
Kenia
3,4 mda
|
KNM-ER
2602
|
||
MAKA
Etiopía
3,4 mda
|
MAK-VP
1/12
|
||
 |
| Australopithecus afarensis. AL 288-1 Lucy |
Morfología.
En Hadar (Etiopía)
se encontraron en 1974 los restos de Lucy
(AL 288-1 datados en 3,2 mda.) de 1 m. de estatura. Con otros restos de Hadar y
Laetoli, se clasificó en la categoría de Australopithecus
afarensis (Johanson, White, Coppens). Los A. afarensis son:
- De acuerdo con
- 1,00-1,65 m. Considerable longitud de las piernas. 20-48 kg (Brassey et al, 2017).
- Capacidad craneal 478 cc (entre 380-540 cc), poco mayor que la de los chimpancés. Cráneo largo y bajo, claramente similar al de un primate, estirado entre el prognatismo alveolar y el ángulo formado por el occipucio. El patrón de crecimiento del temporal es similar al del gorila (Claire E. Terhune et al, 2013). Las superficies de inserción del músculo temporal indican un gran desarrollo de su parte posterior, con poderosa cresta nucal. Asimetría cortical poco marcada. La red de la vena meníngea media es más densa en la parte posterior. Crestas sagital y nucal marcadas. Hueso occipital anguloso entre la placa y el plano nucal. Base del cráneo poco flexionada, acampanada. Orificio occipital en posición relativamente retrasada, pero orientado hacia abajo. Hueso de la parte inferior neumatizado, como en los grandes simios. Gunz et al (2020), a partir del estudio de los endocastos, consideran que la maduración cerebral era dilatada en el tiempo, como en los humanos, al mismo tiempo que la organización cerebral se mantenía similar a la de los chimpancés.
- Prognatismo subnasal. Rostro muy prognato, especialmente en la región alveolar. El plano óseo que une la base de la nariz y la parte del maxilar encima de los incisivos forma un clivus nasoalveolar. Articulación de la mandíbula poco profunda, paladar poco profundo en la parte anterior.
- Arcada dental en U la superior y V intermedia la inferior. No existen pilares óseos verticales que refuercen los márgenes del orificio nasal. La raíz anterior del arco cigomático nace a la altura de M1. Sigue habiendo fosa canina en esta región del maxilar. Arcada dental superior más bien larga, estrecha y poco divergente hacia atrás. Mandíbula robusta, con una sínfisis muy inclinada hacia atrás. Las características dentales nos indican que Australopithecus afarensis está en una rama que se separa de la que dará origen a Homo.
- Incisivos situados lateralmente en relación a la abertura nasal, anchos y con raíces curvas.
- Caninos salientes (especialmente los machos) más pequeños que en los chimpancés y anamensis, pero sustancialmente más grandes que en los humanos. Forma espatulada.
- Los caninos superiores se desgastan por la punta, como en todos los homínidos, pero también en los lados, como en los monos antropomorfos: un pequeño diastema separa el canino inferior del primer premolar inferior P3, para albergar un canino superior saliente.
- La morfología de P3 es intermedia, con una corona asimétrica, pequeña cúspide interna -que puede estar ausente- y otra externa mayor (los chimpancés tienen una sola cúspide y los sapiens dos aproximadamente del mismo tamaño) y amplia variabilidad morfológica.
- El tamaño de los molares aumenta de M1 a M3.
- En las etapas de la emergencia dental es más parecido a los humanos que a los chimpancés (Halszka Glowacka, William H. Kimbel y Donald C. Johanson, 2017).
- Esqueleto postcraneal sin ninguna particularidad con respecto a la
descripción del género Australopithecus: pelvis, fémur, tobillo y pie con características derivadas. Siete vértebras cervicales y sólo 12 vértebras torácicas como los seres humanos, en lugar de 13 como los simios africanos. La transición anatómica entre la parte vertebral torácica y la lumbar se produce en el segmento torácico 11; esta característica es rara en los simios y los seres humanos modernos (Ward et al, 2017). El tórax de Kudanuumuu, con forma alargada, se parece más al del humano
moderno que al de los grandes simios. La coincidencia en la forma del hombro
de Selam y Lucy que muestra la conservación de la adaptación a la trepa ( ver Ruff et al, 2016 para el húmero),
indica que esta característica es propia de la especie y no está
relacionada con la ontogenia. El hombro de Kudanuumuu es más derivado y evidencia una manipulación incipiente. Según
- Para C. Owen Lovejoy afarensis es un bípedo exclusivo. Los rasgos que sugieren un desplazamiento arbóreo serían plesiomorfias indicativas solo de una historia arbórea (Selby y Lovejoy, 2017).
Los seres de Hadar
son una muestra excelente de la evolución en mosaico:
-
Algunos
rasgos, como la cadera, indican una proximidad notable respecto de los
homínidos posteriores.
o
AL
333-160, un cuarto metatarsiano del pie izquierdo datado en 2,3 Ma plano,
similar al humano moderno, mientras que en los grandes simios es curvo.
-
Otros,
como el hombro, la longitud de las extremidades anteriores o el cráneo pequeño
y robusto, son signos de la presencia de caracteres primitivos.
-
Lucy,
AL 333-160 y Kadanuumuu proporcionan la prueba irrefutable de que en el proceso
de evolución primero fue la locomoción y mucho después las capacidades
craneales.
 |
| De izquierda a derecha, arcadas dentales inferiores de ramidus, anamensis y afarensis. |
Modos de vida.
Los hallazgos en los esqueletos muestran que se trataba de una especie que caminaba erguida sin nudilleo pero que también trepaba a los árboles. Lo más probable es que vivieran en pequeños grupos con cierta colaboración entre los individuos (José Enrique Campillo, 2005), durmieran en nidos construidos en los árboles con ramas y en cuevas poco profundas y utilizaran palos y piedras sin manufacturar como instrumentos. Desconocían el uso del fuego. Las débiles crestas sagitales suponen que no tenían un potente aparato masticador, si bien parece que los productos duros y abrasivos iban cobrando importancia.
Hábitat: orillas lacustres con vegetación boscosa y
zonas abiertas en mosaico, lo que sugiere que afarensis exploraba distintos hábitats (Aiello y Andrews, 2000; Denise F. Su y Terry Harrison, 2014). Se trata del mismo hábitat en el que vivió su cronoespecie predecesora, Australopithecus anamensis. A pesar de que el paisaje no había cambiado, la dieta de afarensis aumentó en variedad. Su alimentación, oportunista, estaba compuesta en gran parte por vegetales, pero también por huevos, reptiles, termitas e insectos diversos. La enzima trehalasa está presente en nuestro intestino y sirve para digerir el azúcar trehalosa, abundante en los caparazones de los insectos. Según Wynn, Cerling, Sponheimer et al (2013), Wynn et al (2016), además de hojas y frutas se alimentaba de grandes cantidades de pastos C4 y juncias (un 22% en promedio); también consumía algunas plantas suculentas. En el análisis de microdesgaste realizado por Lucas K. Delezene et al (2013), no se hallaron pruebas de que afarensis procesara alimentos duros o de que la función del P3 inferior cambiase a lo largo del tiempo.
Philip L. Reno et
al (2010) tras estudios de medida de los esqueletos, dedujeron un mayor
dimorfismo sexual que en Ardipithecus. Creen,
junto con Lovejoy, que este aumento del dimorfismo es producto de la selección
sexual. Mientras las hembras escogerían machos más grandes, que garantizaban
mejor defensa y acceso a los recursos en el marco de una expansión de los
australopitecinos con competencia interespecífica, los machos preferirían
hembras más pequeñas, con menor consumo de alimentos. La competencia entre
machos queda descartada debido a que no hay indicios de la misma en Ardipithecus.
 |
| Reconstrucción de Lucy. Brassey et al, 2017. |
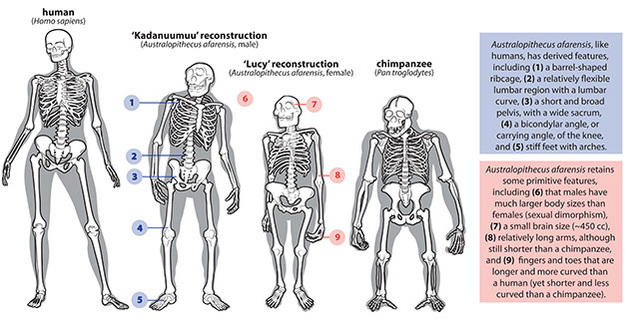 |
| Comparación de Homo sapiens, Australopithecus afarensis y chimpancé. Ward y Hammond, 2016 |
Discusión filogenética
En cuanto a su origen, hasta hace poco, la relación con anamensis parecía clara, pero Haile-Selassie et al (2019), consideran que estas dos especies coexistieron en el tiempo:- BEL-VP-1/1, datado hace 3,9 Ma, e incluso los dientes de Fejej (Ward et al, 2014; 4,2-4 Ma) deberían clasificarse como A. afarensis.
- El cráneo MRD-VP-1/1, hallado en 2016 fue datado de forma segura en hace 3,8 Ma.
- Strait et al (1997) y Grine (2000) proponen que afarensis evolucionó a una especie similar a africanus, con rasgos primitivos, de la que por un lado surgiría africanus como rama muerta y por otro un ancestro común a Homo y Paranthropus. En consecuencia, para conservar la monofila de los géneros, clasifican afarensis dentro de Praeanthropus.
 |
| Posición filogenética de afarensis. Donald Johansson, 2016. |
Cabe preguntarse en qué medida cabe incluir en la misma especie ejemplares de Laetoli y Hadar, muy separados entre sí, además de una distancia en el espacio de 1.800 km., y en el tiempo de 0,8 Ma.
- Tobias (1980) sostuvo sin éxito que todos los ejemplares de Laetoli, Hadar, Sterkfontein y Makapansgar pertenecen al Australopithecus africanus de Dart, aun cuando se puedan identificar subespecies.
- Poirier (1987) opinaba que ver en la muestra de Hadar una o varias especies depende de cuánta variabilidad morfológica debe permitirse dentro de una especie y cuánta variabilidad morfológica puede explicarse por dimorfismos sexuales.
- Conroy (2005) reconoce la posibilidad de dos especies, una de las cuales daría lugar a boisei.
- Olson (1985) atribuyó las diferencias a presiones selectivas.
- Algunos ejemplares habrían desarrollado especializaciones que permitirían clasificarlos como miembros del clado Paranthropus. De este modo, la idea de que la separación entre Paranthropus y Homo ocurrió hace 2,5 Ma, quedaría destruida porque los ejemplares robustos de Hadar tendrían al menos 3 Ma.
- En 1994 Kimbel, Johanson y Rak describieron el ejemplar llamado el hijo de Lucy, de 3 Ma, de cráneo muy ancho pero cuerpo mandibular menos robusto que el promedio. Este ejemplar apoya la hipótesis de una especie única muy variable. Otros hallazgos recientes hablan a favor de unos dimorfismos sexuales grandes.
- White y Johanson han encontrado fósiles con un amplio rango de variación de tamaño en el mismo yacimiento y con una datación en un periodo corto de tiempo. Un fémur largo muestra las mismas adaptaciones que el fémur corto de Lucy.
- En todo el espectro de hábitats se encuentran tanto individuos grandes como pequeños.
- William H. Kimbel (2007) piensa que la especie ofrece un dimorfismo sexual grande junto con una tendencia anagenética al incremento de tamaño del cráneo. Este aumento de tamaño se produce con una gran variedad morfológica propia de una evolución en mosaico. Por otra parte las calvarias de machos jovenes son difícilmente distinguibles de las de hembras adultas.
La opinión más generalizada es considerar una única especie antecesora de todos los homínidos posteriores (Lewin y Foley, 2004; Boyd y Silk, 2004; Harmon, 2006; Donald Johanson, 2016).
Adam van Arsdale: Australopithecus afarensis
Enlaces:
Fossil hominids from the Laetoli Beds. Leakey, M. D., et al. (1976).
A new species of the genus Australopithecus (primates: Hominidae) from the Pliocene of eastern Africa. Johanson, D. C.; White, T. D. y Coppens, Y. (1978).
‘‘Lucy’’ Redux: A Review of Research on Australopithecus afarensis. William H. Kimbel* and Lucas K. Delezene (2009).
Paleoecology and Paleoenvironment: a Case Study of Plio-Pleistocene Mammals from Laetoli. Kris Kovarovic (2012).
Lucy (A. afarensis) and Early Hominid Bi-Pedalism. Melissa Orenstein (2013).
The paleoanthropology of Hadar, Ethiopia. Donald Johanson (2016).
Fossil hominids from the Laetoli Beds. Leakey, M. D., et al. (1976).
A new species of the genus Australopithecus (primates: Hominidae) from the Pliocene of eastern Africa. Johanson, D. C.; White, T. D. y Coppens, Y. (1978).
‘‘Lucy’’ Redux: A Review of Research on Australopithecus afarensis. William H. Kimbel* and Lucas K. Delezene (2009).
Paleoecology and Paleoenvironment: a Case Study of Plio-Pleistocene Mammals from Laetoli. Kris Kovarovic (2012).
Lucy (A. afarensis) and Early Hominid Bi-Pedalism. Melissa Orenstein (2013).
The paleoanthropology of Hadar, Ethiopia. Donald Johanson (2016).
Bibliografía:
- White, Tim D., and Gen Suwa. "Hominid footprints at Laetoli: facts and interpretations." American Journal of Physical Anthropology 72.4 (1987): 485-514.
- Day, M. H., and E. H. Wickens. "Laetoli pliocene hominid footprints and bipedalism." Nature (1980).
- Leakey, Mary Douglas, and John Michael Harris. Laetoli, a Pliocene site in northern Tanzania. Clarendon Press. Oxford University Press, 1987.
- White, Tim D. "Additional fossil hominids from Laetoli, Tanzania: 1976–1979 specimens." American Journal of Physical Anthropology 53.4 (1980): 487-504.
- Raichlen, David A., et al. "Laetoli footprints preserve earliest direct evidence of human-like bipedal biomechanics." PLoS One 5.3 (2010): e9769.
- Stern Jr, Jack T., and Randall L. Susman. "The locomotor anatomy of Australopithecus afarensis." American Journal of Physical Anthropology 60.3 (1983): 279-317.
- Kimbel, William H., et al. The skull of Australopithecus afarensis. Oxford University Press, 2004.
- Ward, Carol V. "Interpreting the posture and locomotion of Australopithecus afarensis: Where do we stand?." American journal of physical anthropology119.S35 (2002): 185-215.
- Johanson, Donald C., et al. "Morphology of the Pliocene partial hominid skeleton (AL 288‐1) from the Hadar formation, Ethiopia." American Journal of Physical Anthropology 57.4 (1982): 403-451.
- Alemseged, Zeresenay, et al. "A juvenile early hominin skeleton from Dikika, Ethiopia." Nature 443.7109 (2006): 296-301.
- McPherron, Shannon P., et al. "Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia." Nature466.7308 (2010): 857-860.
- Wynn, Jonathan G., et al. "Geological and palaeontological context of a Pliocene juvenile hominin at Dikika, Ethiopia." Nature 443.7109 (2006): 332-336.
Te puede interesar:
|