Principales hallazgos de australopitecos y parántropos. Bacon, 2000. En Malapa, a 15 km de Swartkrans y Kromdraai, apareció en 2010 A. sediba.
Australopithecus se definió originalmente por Raymond Dart en 1925 sobre la base del cráneo del Niño de Taung, hallado en Sudáfrica, con una datación de 3-2 Ma, con un pequeño tamaño, pero con dientes delanteros más pequeños que los del chimpancé, incluyendo los caninos, y unos molares más grandes. Lo más importante es la posición del foramen magnum que sugiere una bipedación. Dart nombró a esta especie Australopithecus africanus (Dart 1925).
Desde entonces, se han descubierto varias especies más. Los australopitecinos más antiguos y primitivos aparecieron en el este de África, en particular Etiopía y Kenia, y en Sudáfrica los más derivados. Los fósiles de África Oriental son a menudo fragmentarios porque debido a la actividad geológica o de los carnívoros, pero a cambio las dataciones son fiables. Los ejemplares sudafricanos, por el contrario, están bien conservados pero las dataciones no son seguras (Ward y Hammond, 2016)
Hay buena evidencia de que A. anamensis y A. afarensis representan cronoespecies dentro de un amplio linaje (Kimbel et al, 2006), pero las relaciones con las otras especies del género no son claras. Puede ser que el género esté formado por una amalgama de especies que no están estrechamente relacionadas (grupo parafilético), a pesar de que parecen compartir un común grado de adaptación:
Hominini bípedos que conservan la suspensión y la capacidad de trepa (4,5-2,6 Ma) de rostro robusto y cerebro relativamente desarrollado. 1,05-1,45 m. 27-60 kg
Aguirre (1970) indica que la distinción entre Australopithecus y Paranthropus no es fácil.
Para Adam Gordon (2013) el dimorfismo sexual en Australopithecus fue más bien resultado de la selección natural, que de la selección sexual. Este sería el grado de dimorfismo por especie:
El dimorfismo en Ardipithecus ramidus es difícil de asegurar.
Los hallazgos de Australopithecus anamensis son demasiado escasos para poder estimar su dimorfismo.
Australopithecus afarensis, probablemente tenía un dimorfismo algo mayor que el de los chimpancés y bonobos.
Autralopithecus africanus, algo menor que el de los chimpancés y bonobos.
Australopithecus sediba mostraba un dimorfismo muy pequeño.
Morfología craneal de Australopithecus comparada con Proconsul y Pan
Masa y altura de australopitecinos
Características comparadas de Australopithecus y Paranthropus
Cráneo
El foramen magnum ocupa una posición avanzada en la base del cráneo orientada hacia abajo, como en el hombre. Menor desarrollo de los músculos de la nuca que en el chimpancé. Cresta nucal transversal en posición inferior.
Caja craneana más desarrollada y más elevada en el parietal que en el chimpancé, con rostro menos prognato y más ancho. Huesos de la bóveda craneal más bien delgados. Continuidad entre la región supraorbital y la placa del frontal. En su parte posterior, el hueso occipital se divide en una placa de perfil redondeado más amplia que el plano nucal, orientado hacia abajo y hacia delante. Las dos partes del occipital están separadas por una cresta transversal que en los machos se une a las líneas temporales para formar una cresta temporal-nucal.
El cerebro muestra el comienzo de tendencias evolutivas. Volumen cerebral 380-500 cc, mayor que el de los chimpancés (350-400). Mayor coeficiente de encefalización (2,6-2,3 frente a 2,1). Áreas corticales, sobre todo la parietal, más grandes, ligeramente asimétricas. La red sanguínea asociada a la vena meníngea media, indica una irrigación más densa en la parte parietal.
Organización cerebral similar a la del chimpancé. Sin embargo, la maduración cerebral se alarga en el tiempo, como en los humanos (Gunz et al, 2020).
Rostro alto y poderoso. El avance de las arcadas dentales se manifiesta en un prognatismo alveolar, que solo moviliza la parte alveolar del hueso maxilar. El prognatismo de las partes media y superior del rostro es acusado. Paladar largo y poco profundo. Unos refuerzos óseos consolidan los márgenes del orificio nasal (orificio piriforme). La raíz anterior del arco cigomático, que forma la apófisis piramidal del maxilar, nace encima del primer molar o del segundo premolar. Esta disposición propicia la llamada fosa canina. La región situada encima de las órbitas, forma un torus continuo, sin depresión en la glabela. Vistas de frente, las líneas temporales anteriores convergen rápidamente hacia la línea sagital de la bóveda craneal. Vista desde arriba, la constricción entre las órbitas y la caja craneana es marcada.
Mandíbula relativamente corta, con una rama ascendente muy desarrollada. Las marcas de inserción de los músculos masticadores son profundas. La parte anterior y horizontal de la mandíbula es alta y ancha, con una robustez asegurada por la presencia de varios repliegues óseos, sobre todo en la sínfisis. Los torus transversales inferior y superior consolidan la sínfisis en su parte interna. Ausencia de mentón.
Los dientes forman arcadas dentales divergentes hacia atrás en forma de V o U de ramas más o menos abiertas (parabólicas en bahrelghazali, en forma de U en anamensis, en forma de V en afarensis). Masticación potente, más que la de los chimpancés y los humanos.
Los incisivos son cortos y robustos. La corona es ancha.
Los caninos son prominentes y su desgaste se efectúa desde la punta hacia la raíz.
Premolares robustos con coronas formadas por dos cúspides desarrolladas.
Molares grandes, con coronas anchas en sentido bucolingual. Tamaño creciente de M1 a M3.
Raíces de premolares y molares largas, robustas y profundamente implantadas en los alveolos del maxilar y la mandíbula.
Esmalte grueso.
Una raíz del cigomático más anteriormente posicionada mejoró la eficiencia de la fuerza de mordida mientras que otras características faciales fueron seleccionadas para compensar el debilitamiento del rostro (Ledogar et al, 2016)
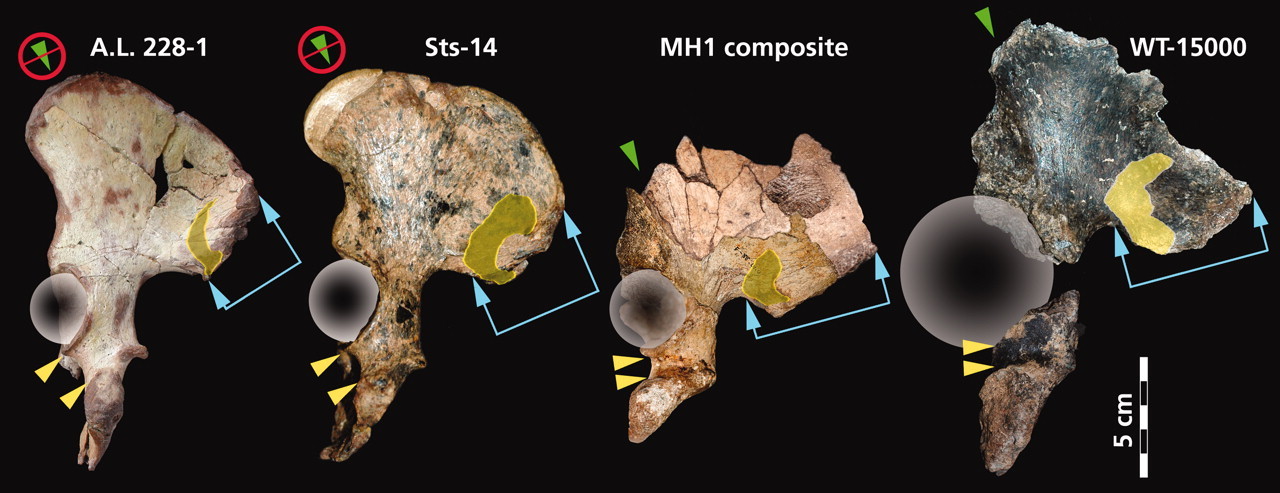
Comparación de los coxis de Au. afarensis (AL 288-1), Au. africanus (Sts 14), Au. sediba (MH1), y H. erectus (KNM-WT 15000). Science.
Restos postcraneales.
La parte superior del cuerpo se parece a la de los chimpancés o bonobos, mientras la inferior es más parecida a la del humano.
Tronco
Cuerpo más bien ancho y achaparrado. Tronco en forma de cono, como el de los chimpancés: caja torácica muy ancha en la parte interior que va estrechándose conforme se acerca al cuello. Espina curvada.
Posición erguida.
El homóplato, la clavícula y los tubérculos óseos que forman la articulación del hombro, son robustos y orientados hacia arriba. Las inserciones musculares denotan una musculatura potente en el cinturón escapular.
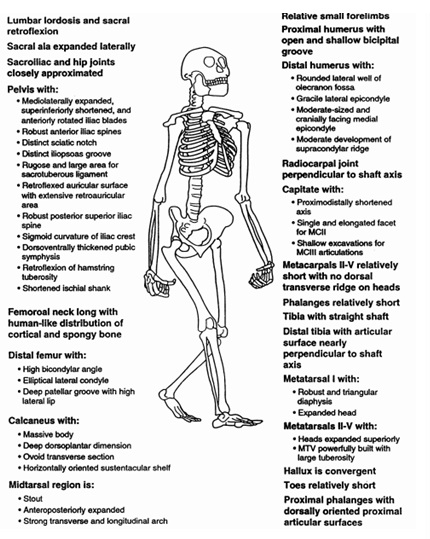
Pelvis tan ancha como en los humanos actuales, menos abierta hacia delante y más abierta a los lados y acampanada en los costados, corta y en forma de cubeta, apta para la bipedación. Sacro corto y ensanchado, aunque menos que en el hombre; se inserta como una moneda entre las dos alas ilíacas, asegurando la transmisión del peso del cuerpo. Distancia corta entre la articulación sacro ilíaca y la del fémur, con acetábulo reforzado en sus márgenes, de forma distinta a chimpancés y humanos. Cabeza del fémur poco insertada en el acetábulo. Trabéculos y pilares óseos refuerzan esta región. Inserciones ligamentosas menos fuertes que en el hombre. El perfil de las alas ilíacas sigue una línea recta desde el sacro hasta la espina ilíaca anterior. Huesos púbicos largos, con sínfisis púbica relativamente esbelta. Las características de la pelvis indican que estos seres eran bípedos.
Últimas costillas situadas muy cerca del borde superior de la pelvis, lo que revela la práctica ausencia de talle. 6 vértebras lumbares, como en los monos antropomorfos (5 en los hombres), si bien presenta lordosis.
Extremidades superiores
Brazos largos.
La fosa de la base del húmero, que forma la articulación del codo (olecraneana) es profunda, como en los chimpancés, y permite el bloqueo del codo durante la suspensión o la braquiación.
Morfología distal diafisaria del húmero (Lague, 2015)
La presente evidencia indica que la forma diafisaria de Australopithecus se caracteriza por un patrón de estabilidad a largo plazo. Entre las especies de Australopithecus, sólo es inusual en Au. sediba. Además, la morfología diafisaria distal de P. robustus se asemeja a la de los Australopithecus no-sediba. Este patrón homogéneo a largo plazo entre los australopitecinos no es exclusivo de la diáfisis humeral, ya que también se ha observado para la morfología articular del codo (Lague y Jungers, 1996; Lague, 2014) y para la morfología femoral proximal (Richmond y Jungers, 2008; Richmond y Jungers, 2012). Vale la pena señalar, sin embargo, que aunque el fémur proximal de P. boisei no parece diferir considerablemente del de otros australopitecinos (basado en KNM-ER 1504), el húmero distal de P. boisei se aparta de la pauta general para los australopitecinos tanto en la morfología articular como en la forma de la sección transversal diafisaria. Con respecto a la morfología distal diafisaria del húmero, las similitudes generales observadas entre Australopithecus (no sediba), P. robustus, y múltiples Hominini, sugieren que la morfología de los Australopithecus no sediba representa una condición primitiva. La marcada variación en la morfología diafisaria observada entre las especies posteriores a Australopithecus puede reflejar un aumento de la diversidad de uso de la extremidad anterior en el Pleistoceno Inferior, quizás asociada a la heterogeneidad más pronunciada en el hábitat y los recursos (Antón, Potts y Aiello, 2014).
Los huesos del antebrazo son largos y curvados.
Los huesos de la mano recuerdan los de los monos antropomorfos arborícolas: los del carpo robustos, los del metacarpo largos, primeras falanges largas y curvadas, últimas falanges estrechas. No se observa ningún carácter que recuerde el nudilleo de los monos antropomorfos africanos actuales. Pulgar largo y estrecho en relación a la mano, no oponible ni capaz de presa de martillo.
Combinación en el hombro de rasgos primitivos y derivados. El comportamiento arbóreo que se manifiesta en el húmero proximal y está relacionado principalmente con la posición sostenida por debajo de la rama (Julia Arias-Martorell et al, 2015).
Extremidades inferiores
Piernas cortas.
Cuello del fémur largo, más bien esbelto. La diáfisis forma un ángulo con la vertical de forma que el eje del fémur converge hacia las rodillas (valgo).
Articulación de la rodilla más flexible que en el humano. Mantenimiento de la rodilla en posición flexionada en el curso de la marcha.
La meseta tibial ligeramente convexa y el cóndilo lateral de fémur, denotan un perfil más circular. Christopher B. Ruff y Ryan Higgins (2013) han analizado la morfología femoral proximal. Las observaciones son consistentes con un patrón de marcha ligeramente alterada, con una desviación lateral mayor del centro de masa del cuerpo.
El pie recuerda el de los monos antropomorfos arborícolas. Apoyo del pie sobre su borde lateral durante la marcha, en varus. Tarso, metatarso y falange de igual longitud. Metatarso y falanges relativamente más largas que en los chimpancés. El tarso permite movimientos variados, con huesos largos pero sin bóveda plantar. Primera falanges largas y curvadas. Dedo gordo completamente aducido como el del sapiens.
Resumen:
Poco más altos que los chimpancés.
Capacidad craneal: 500-380 cc.
Bipedia similar a la de los humanos modernos.
África del Este y Sudáfrica.
Enorme variación en el tiempo 4-1 mda (si incluimos los parántropos).
Rasgos primitivos
Cráneo alargado.
Caja aplanada.
Frente huidiza.
Torus supraorbital muy marcado.
Prognatismo.
Ausencia de mentón.
Conservación de adaptaciones para la trepa en las extremidades superiores.
Rasgos derivados
Arco dentario en forma parabólica.
Relacionados con la bipedia.
Posición horizontal del foramen magnum.
Columna curvada.
Pelvis corta.
Amplia zona de inserción de los músculos con los glúteos.
Hallux aducido.
Algunos artefactos líticos, han sido atribuidos a los australopitecinos.
Se han hallado huesos con marcas de corte en Dikika, Etiopía, con una datación de 3,4 Ma, donde solo se conocen restos de A. afarensis (McPherron et al, 2010).
La industria Lomekwiense (Kenia) se ha datado en 3,3 Ma (Harmand et al, 2015), en el mismo contexto geográfico y cronológico de Kenyanthropus platyops y contemporáneas de A. afarensis.
La mayoría de la evidencia actual sugiere que los australopitecos vivieron una variedad de entornos heterogéneos, incluyendo bosques secos y húmedos, matorrales, bosques de ribera, márgenes de los lagos y praderas arboladas (Behrensmeyer y Reed, 2013).
Distribución geográfica de los australopitecos, a lo largo de las cuencas africanas. Las líneas indican las rutas de conexión, con fechas en millones de años. Robert A. Foley, 2013.
Cladograma de los australopitecineos, según Strait y Grine (2004)
Tres visiones diferentes de la evolución de los australopitecos; a) anagénesis; b) cladogénesis; c) alopatria. Robert A. Foley, 2013.
Importancia en la evolución homínida
Con el sucesivo descubrimiento de los yacimientos sudáfricanos fue imponiéndose la tesis dispersora de Broom y se ha tendido a clasificar los restos según el siguiente esquema:
Australopithecus convencionalmente “grácil” por comparación. Las proporciones más armoniosas entre los dientes anteriores y los molares indican un régimen más omnívoro.
La morfología de la cara y los dientes, sugiere que los australopitecinos se adaptaron para consumir alimentos duros, como tubérculos, frutos secos, semillas o raíces, al menos durante las épocas de escasez de alimentos. Los estudios de isótopos, de desgaste y de partículas, parecen confirmar una dieta dura (Henry et al, 2012).
Sin embargo, según Frederick Grine et al (2013) todos los fósiles molares de afarensis y anamensis caen dentro del rango de microdesgaste de Gorilla gorilla. Esto sugiere el mantenimiento de una dieta de hojas blandas aunque la dentadura fuese lo suficientemente potente como para recurrir a alimentos duros en caso de necesidad.
No sabemos como se produce la separación en estos dos grupos más especializados, pero para Lockwood y Tobias A. africanus es la especie ancestral de:
Una forma robusta especializada en una alimentación herbívora dura, Paranthropus.
Homo, de forma grácil, asociado a artefactos primitivos.
Robert A. Foley (2013) ha investigado hasta qué punto los modelos evolucionistas arrojan alguna luz a la especiación de los australopitecos.
El modelo de radiación adaptativa parapátrida no parece aplicable. La evolución de los homínidos en general y la de los australopitecos en particular no muestra una especiación explosiva sino más bien una dispersión direccional sencilla no asociada a los altos niveles de selección sexual frecuentes en la radiación.
Los modelos alopátridos son consistentes con la dispersión geográfica de las especies del género. Esta dispersión, aparece como la fuerza conductora de la especiación. Debido a que en toda su distribución geográfica los australopitecos ocuparon hábitats muy similares, su nivel de diversificación y especiación fue moderado.
Sinapomorfias humanas en Australopithecus
Adam van Arsdale: Australopithecus hallados en Sudáfrica
Aiello, L., & Dean, C. An Introduction to Human Evolutionary Anatomy. London: Academic Press (1990).
Alemseged, Z., Spoor, F., et al. A juvenile early hominin skeleton from Dikika, Ethiopia. Nature, 443, 296-301 (2006).
Asfaw, B., White, T., et al. Australopithecus garhi: a new species of early hominid from Ethiopia. Science, 284, 629-635 (1999).
Behrensmeyer, A., & Reed, K. Reconstructing the habitats of Australopithecus: paleoenvionments, site taphonomy, and faunas. The Paleobiology of Australopithecus. ED: Reed K.E., Fleagle J.G., Leakey R. E. New York: Springer. 41-60 (2013).
Berger, L. R., de Ruiter, D. J., et al. Australopithecus sediba: a new species of Homo-like australopith from South Africa. Science, 328, 195-204 (2010).
Brown, B., Brown, F. H., et al. New hominids from the Lake Turkana Basin, Kenya. Journal of Human Evolution, 41, 29-44 (2001).
Brunet, M., Beauvilain, A., et al. The first australopithecine 2,500 kilometres west of the Rift Valley (Chad). Nature, 378, 273-275 (1995).
Coffing, K., Feibel, C., et al. Four-million-year-old hominids from East Lake Turkana, Kenya. American Journal of Physical Anthropology, 93, 55-65 (1994).
Dart, R. A. Australopithecus africanus: the man-ape of South Africa. Nature, 115, 195 (1925).
Falk, D., Redmond, J. C., et al. Early hominid brain evolution: a new look at old endocasts. Journal of Human Evolution, 38, 695-717 (2000).
Fleagle, J. G., Rasmussen, D. T., et al. New hominid fossils from Fejej, Southern Ethiopia. J. Hum. Evol., 21, 145-152 (1991).
Haile-Selassie, Y., Latimer, B. M., et al. An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia. Proc Natl Acad Sci U S A, 107, 12121-12126 (2010a).
Haile-Selassie, Y., Saylor, B. Z., et al. New Hominid Fossils From Woranso-Mille (Central Afar, Ethiopia) and Taxonomy of Early Australopithecus. American Journal of Physical Anthropology, 141, 406-417 (2010b).
Haile-Selassie, Y., Gibert, L., et al. New species from Ethiopia further expands Middle Pliocene hominin diversity. Nature, 521, 483-488.
Harmand, S., Lewis, J. E., et al. 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521, 310-315.
Hartwig-Scherer, S. Body weight prediction in early fossil hominids: towards a taxon-"independent" approach. American Journal of Physical Anthropology, 92, 17-36 (1993).
Henry, A. G., Ungar, P. S., et al. The diet of Australopithecus sediba. Nature, 487, 90-93 (2012).
Holloway, R. L. (1975) Early hominid endocasts: volumes, morphology, and significance for hominid evolution. In Tuttle, R. H. (Ed.), Primate Functional Morphology and Evolution. Paris: Mouton.
Johanson, D. C., Lovejoy, C. O., et al. Morphology of the Pliocene partial hominid skeleton (A. L. 288-1) from the Hadar formation, Ethiopia. American Journal of Physical Anthropology, 57, 403-452 (1982a).
Johanson, D. C., & Taieb, M. Pliocene hominid discoveries in Hadar, Ethiopia. Nature, 260, 293-297 (1976).
Johanson, D. C., Taieb, M., et al. Pliocene hominids from the Hadar formation, Ethiopia (1973-1977): stratigraphic, chronologic and paleoenvironmental contexts, with notes on hominid morphology and systematics. American Journal of Physical Anthropology, 57, 373-402 (1982b).
Johanson, D. C., White, T. D., et al. A new species of the genus Australopithecus (Primates: Hominidae) from the Pliocene of eastern Africa. Kirtlandia, 28, 1-14 (1978).
Kimbel, W. H. Identification of a partial cranium of Australopithecus afarensis from Koobi Fora Formation, Kenya. Journal of Human Evolution, 17, 647-656 (1988).
Kimbel, W. H., & Delezene, L. K. "Lucy" redux: a review of research on Australopithecus afarensis. Am J Phys Anthropol, 140 Suppl 49, 2-48 (2009).
Kimbel, W. H., Johanson, D. C., et al. The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia. Nature, 368, 449-451 (1994).
Kimbel, W. H., Lockwood, C. A., et al. Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. J Hum Evol, 51, 134-152 (2006).
Latimer, B. (1991) Locomotor adaptations in Australopithecus afarensis: the issue of arboreality. In Coppens, Y. & Senut, B. (Eds.), Origine(s) de la Bipédie chez les Hominidés (pp. 169-176). Paris: Centre National de la Recherche Scientifique.
Leakey, M. Discoveries at Laetoli in northern Tanzania. Proceedings. Geologists Association., 92, 81-86 (1981).
Leakey, M. G., Spoor, F., et al. New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature, 410, 433-440 (2001).
Lockwood, C. A. Sexual dimorphism in the face of Australopithecus africanus. American Journal of Physical Anthropology, 108, 97-127 (1999).
Lockwood, C. A., Richmond, B. G., et al. Randomization procedures and sexual dimorphism in Australopithecus afarensis. Journal of Human Evolution, 31, 537-548 (1996).
McDougall, I., & Brown, F. Geochronology of the pre-KBS Tuff sequence, Omo Group, Turkana Basin. Journal of the Geological Society, 165, 549-562 (2008).
McHenry, H. M. Body size and proportions in early hominids. American Journal of Physical Anthropology, 87, 407-431 (1992).
McPherron, S. P., Alemseged, Z., et al. Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ato at Dikika, Ethiopia. Nature, 466, 857-860.
Plavcan, J. M., Lockwood, C. A., et al. Sexual dimorphism in Australopithecus afarensis revisited: how strong is the case for a human-like pattern of dimorphism? Journal of Human Evolution, 48, 313-320 (2005).
Reno, P. L., Meindl, R. S., et al. Sexual dimorphism in Australopithecus afarensis was similar to that of modern humans. Proc Natl Acad Sci U S A, 100, 9404-9409 (2003).
Reno, P. L., Meindl, R. S., et al. The case is unchanged and remains robust: Australopithecus afarensis exhibits only moderate skeletal dimorphism. Journal of Human Evolution, 49, 279-288 (2005).
Spoor, F., Leakey, M. G., et al. Hominin diversity in the Middle Pliocene of eastern Africa: the maxilla of KNM-WT 40000. Philosophical Transactions B, 365, 3377-3388 (2010).
Stern, J. T. Climbing to the top: a personal memoir of Australopithecus afarensis. Evolutionary Anthropology, 9, 113-133 (2000).
Tobias, P. V. (1975) Brain evolution in the Hominoidea. In Tuttle, R. H. (Ed.), Primate Functional Morphology and Evolution. Paris: Mouton.
Ward, C., Kimbel, W. H., et al. Complete Fourth Metatarsal and Arches in the Foot of Australopithecus afarensis. Science, 331, 750-753 (2011).
Ward, C. V. Interpreting the posture and locomotion of Australopithecus afarensis: where do we stand? American Journal of Physical Anthropology, Suppl 35, 185-215 (2002).
Ward, C. V., Leakey, M. G., et al. Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. Journal of Human Evolution, 41, 255-368 (2001).
White, T. D., Suwa, G., et al. Jaws and teeth of Australopithecus afarensis from Maka, Middle Awash, Ethiopia. American Journal of Physical Anthropology, 111, 45-68 (2000).
White, T. D., WoldeGabriel, G., et al. Asa Issie, Aramis and the origin of Australopithecus. Nature, 440, 883-889 (2006).








crean en dios
ResponderEliminarGran información sobre los peronistas!
ResponderEliminarLos peronchorristas todos unidos chorrearemos
ResponderEliminarDios no existe!!!
ResponderEliminarTu eres quien insulto a dios
ResponderEliminarresponde maldito
ResponderEliminar