 |
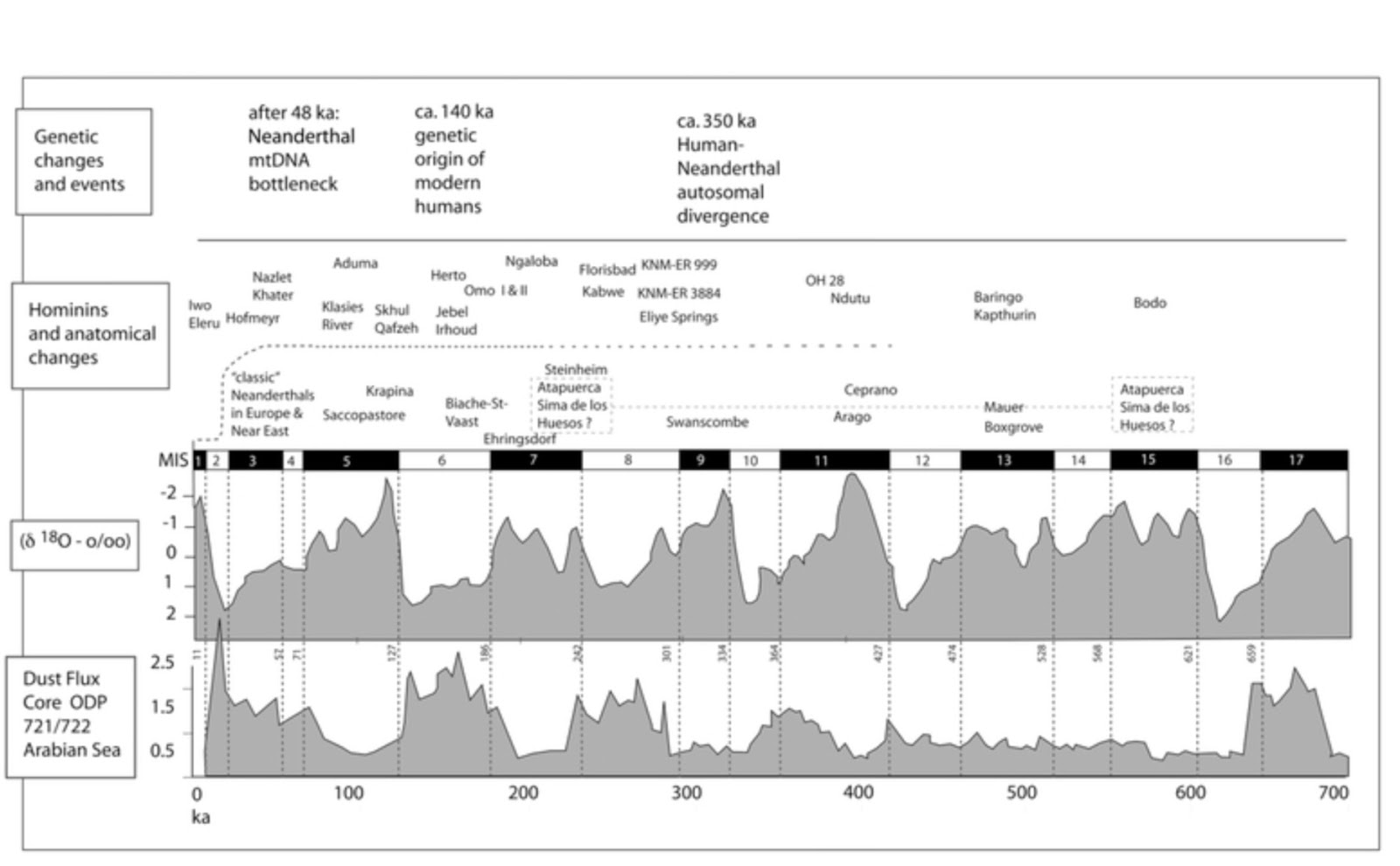
| Principales acontecimientos en la evolución genética, especímenes fósiles, curvas MIS (Klein, 2009) y flujo de polvo desde el Mar Arábigo (Donges et al). |
Al final del Paleolítico Medio, los neandertales y los HAM se diferenciaban claramente en su morfología y tal vez en su comportamiento. En una revisión de 75 características craneales, dentales y postcraneanas de los neandertales y los humanos modernos tempranos, Trinkaus (2006) llegó a la conclusión de que sólo una cuarta parte eran exclusivas de los neandertales, mientras que muchas más eran exclusivas de los humanos modernos, un hallazgo que implica que la morfología neandertal se había mantenido bastante primitiva, mientras que la de los modernos tempranos era mucho más derivada. Esto podría proporcionar evidencia de que los primeros humanos modernos se habían desplazado a nuevos nichos experimentando importante selección para adaptarse a nuevos hábitos.
- En Europa, las apomorfias del linaje Neandertal incluyeron prognatismo mediofacial, posición posterior del foramen mentoniano, espacio retromolar, amplia fosa suprainíaca de forma ovalada, gran proceso juxtamastoideo junto con un pequeño proceso mastoideo, moño occipital, arcos ciliares de doble arco que se reducen en volumen absoluto y grosor vertical en comparación con los de los homínidos del Pleistoceno Medio, y un cerebro considerablemente más grande que el de la mayoría de los homínidos del Pleistoceno Medio (Hublin 2009 ; Stringer 2007). Este conjunto de características era común entre los homínidos europeos de MIS 5, incluyendo los especímenes de Krapina y Saccopatore, y se hizo aún más frecuente en MIS 4-3. Este aumento gradual en la similitud de los últimos Neandertales se ha denominado el "modelo de acreción" y se desarrolló como una tendencia a largo o tal vez en dos pulsos (Hublin 2009). Un patrón similar se aplica a la evolución de la morfología postcraneal (Trinkaus, 1983, 2006). Los escenarios para la evolución de la morfología postcraneal neandertal han tendido a enfatizar la necesidad de producir grandes cantidades de fuerza física o de conservar el calor en un clima frío.
- Los fósiles de la Sima de los Huesos muestran características en mosaico en prácticamente todos los aspectos de su morfología (Arsuaga et al, 1997). Martinón-Torres et al (2012) han demostrado que algunas de las características de los dientes no métricas son aún más comunes en la muestra de la Sima de los Huesos, que en los neandertales tardíos "clásicos" MIS 4-3, lo que arroja dudas sobre modelos evolutivos lineales de incremento constante.
- Hipótesis alternativas recientes aceptan una datación más antigua en los restos de la Sima de los Huesos y proponen la existencia de dos linajes en Europa hasta 300-400 ka (García y Arsuaga 2011) o complicados escenarios de extinción local, recolonización y mezcla de dos o más poblaciones ( Dennell, Martinón-Torres, y Bermúdez de Castro 2011).
- La evidencia de que los cuerpos neandertales estaban adaptados a un clima frío está en sus caderas anchas, los segmentos distales de las extremidades más cortas, extremidades cortas con relación a la longitud del tronco, grandes superficies articulares y gruesos huesos largos, que también caracterizan a los seres humanos recientes, cuyos antepasados han vivido en climas fríos durante miles de años (Holliday 1997; Pearson 2000; Ruff 1994). La caderas anchas y los huesos largos robustos ya estaban presentes en la muestra de la Sima de los Huesos (Arsuaga et al. 1999;. Bonmatí et al 2010) y pueden haber sido la condición primitiva de Pleistoceno Medio (Arsuaga et al, 1999). Las caderas anchas también pueden haber sido heredadas de Homo erectus (Simpson et al. 2008).
- Recientemente, Betti, von Cramon-Taubadel, y Lycett (2012) demostraron que las dimensiones de la pelvis humana está más relacionada con la historia de la población (distancia de África) que con el clima mientras que las dimensiones del fémur y la tibia correlacionan con la temperatura mínima anual. Esto implica que la forma de la pelvis parece haber seguido un patrón de evolución en gran medida neutral como la mayoría de las dimensiones craneales humanas (Betti et al, 2009, 2010; Roseman 2004; Weaver, Roseman, y Stringer 2007 , 2008). Los segmentos distales de las extremidades en la muestra de la Sima de los Huesos parecen menos acortados que en los neandertales (Carretero et al 2012), lo que proporciona cierta evidencia de que los homínidos europeos evolucionaron para adaptarse al frío.
- En África, los homínidos evolucionaron gradualmente hacia una forma moderna (Bräuer 2008; Pearson 2008; Rightmire 2009). Las apomorfias incluyen un cerebro más grande, un cráneo más globular (Lieberman 2011; Lieberman, McBratney y Krovitz 2002) y un lóbulo temporal ampliado (Lieberman, 2011); una trayectoria de crecimiento alterado endocraneal en relación con los neandertales (Gunz et al. 2012); una cara vertical corta bajo el lóbulo frontal (Lieberman 2011); retención de una fosa canina en la edad adulta, y la presencia de una barbilla que sobresale en la mandíbula (Stringer 2002, 2007). Los primeros cráneos que muestran un conjunto completo de características modernas son Omo I, datado en 195 ka (Brown, McDougall y Fleagle 2012 ; Día y Stringer 1991; McDougall, Brown, y Fleagle 2005), y los cráneos de Herto de 150-160 ka (Clark et al. 2003; White et al. 2003).
- Los avances en la formación de imágenes, han revelado que los niños neandertales maduraban más rápidamente que los niños modernos (Smith et al, 2010). La dentición mandibular de Jebel Irhoud 3, un homínido de 160 ka con afinidades a los humanos modernos (Hublin 2001; Hublin y Tillier 1981), conserva la evidencia de un ritmo más lento en el desarrollo dental (Smith et al, 2007). Por tanto, los neandertales estaban más cerca de la condición ancestral de una más rápida maduración esquelética dental de Homo erectus (Dean y Smith 2009; Dean et al. 2001; Graves et al 2010).
- Europeos y africanos del Pleistoceno Medio tienden a ser de alta a mediana estatura (Carretero et al. 2012) y muy pesados para su altura en relación con los cazadores y recolectores modernos (Churchill et al, 2012; Kappelman 1996). Se observa un físico más esbelto en Omo I (Pearson et al, 2008) y en los primeros humanos modernos de Skhul y Qafzeh en Israel (Carretero et al, 2012; Ruff, Trinkaus y Holliday 1997). Los neandertales eran al menos un 20% más pesados que los forrajeros humanos modernos de similar altura (Kappelman 1996 ; Ruff, Trinkaus y Holliday 1997). Carretero et al (2012) propusieron que esta reducción de la masa corporal puede haber sido una adaptación evolutiva a un estilo de vida que favorece la eficiencia energética. La estatura de los primeros humanos modernos durante el Levallois-Musteriense del Levante Mediterráneo y el Gravetiense de Europa es particularmente sorprendente en relación con la de los neandertales y con casi todas las otras muestras europeas de antes del siglo XX (Carretero et al, 2012).
Se han explicado las diferencias en base a una selección direccional para el clima, pero no se puede descartar la alternativa de la deriva genética (Howell 1957). Los dos procesos no son mutuamente excluyentes, y ambos actúan a menudo al mismo tiempo.
Según Osbjorn M. Pearson, los datos ambientales y genéticos sugieren que en los homínidos europeos (neandertales) el principal factor evolutivo fue la deriva, mientras que en África (HAM) operaron tanto la deriva como la selección.
- La deriva genética, se acelera durante los períodos de baja población. Las mutaciones favorables tienden a perderse a causa de la deriva. Weaver, Roseman, y Stringer (2007, 2008) demostraron que si se aplica un modelo de evolución neutral a las dimensiones craneales, la divergencia morfológica observada entre los seres humanos y los neandertales podría explicarse como el resultado de la deriva genética en los últimos 350 ka. Algunos métodos recientes hacen hincapié en el papel del azar, sobre todo si el cambio depende del tamaño de la población y la densidad (por ejemplo, Powell, Shennan, y Thomas, 2009; Shennan 2001).
- Probablemente a causa de los ciclos climáticos, la población euroasiática (neandertales, denisovanos y HAM) fue muy baja durante miles de años.
- Los neandertales, bordearon la extinción durante la mayor parte de su existencia, con un importante desplome de población en MIS 4.
- Los nuevos datos del genoma denisovano (Meyer et al 2012) sugieren que esta población también tenía una sorprendentemente bajo tamaño efectivo de la población a largo plazo, de unos
individuos para el período comprendido entre 400 y 100 ka (Li, Patterson, y Reich, 2012 ).
- La inestabilidad climática afectó también al tamaño efectivo de la poblaciones ancestrales de los modernos europeos y de los chinos, que experimentaron uno o más graves cuellos de botella entre hace 40 y 20 ka tales que el tamaño efectivo de cada una de estas poblaciones se redujo aproximadamente a
antes de repuntar a un tamaño mayor durante el Holoceno (Li y Durbin 2011).
- En África, el efecto de los cambios climáticos sobre la población humana es claramente evidente en la disminución dramática del número de hallazgos correspondientes al Último Máximo Glacial en África Oriental (Brooks y Robertshaw, 1990). Las dificultades para la población humana probablemente continuaron hasta ca. 14,5 ka, como indica la desecación del lago Victoria (Williams et al, 2006) y la desecación del lago Tana (Lamb et al, 2007; Marshall et al 2011).
- La selección, opera mejor cuando las poblaciones son grandes y en expansión; en esas condiciones podemos esperar un mayor número de mutaciones favorables (Cochran y Harpending 2009 ; Hawks et al, 2007). La selección trabaja generalmente en un gen sólo si existen diferentes alelos y uno confiere mayor aptitud que otra, aunque la epistasis (la interdependencia de los genes para producir un fenotipo) puede producir un desplazamiento de objetivo para la selección. La última fuente de nuevos alelos es la mutación. La mayoría de las mutaciones son o bien neutras (y no tienen efecto en la selección natural) o perjudiciales (por interferir con la función del gen y por lo tanto causar efectos perjudiciales para el organismo); sólo un pequeño número de mutaciones llegar a ser beneficioso. Los alelos bajo fuerte selección positiva se mueven rápidamente a la fijación, mientras que los alelos bajo fuerte selección negativa se eliminan rápidamente de una población (Futuyma, 1986).
- Si la selección fue el factor crucial, los principales cambios en la anatomía de cada linaje deben emerger durante los períodos de población de gran tamaño. Si el factor crucial fue la deriva, las nuevas morfologías deben aparecer durante o inmediatamente después de periodos de bajo número de población. Cualquier prueba de estas hipótesis se enfrenta a limitaciones prácticas, incluyendo el registro incompleto de fósiles, los problemas de datación, la resolución inadecuada de los métodos actuales en la localización de los cambios morfológicos o genéticos y la variabilidad de los los registros de flujo de polvo en África, diferentes para cada zona.
- La edad de los fósiles de la Sima de los Huesos es un problema clave para dar sentido al ritmo y modo de evolución de los homínidos en Europa en los últimos 500 ka (Stringer 2012). Si la Sima de los Huesos se remonta a 500-600 ka (Bischoff et al. 2007), se puede pensar que los rasgos neandertales "clásicos" aumentaron en frecuencia muy lentamente dentro del linaje neandertal. Una datación corta (350 ka; Stringer, 2012), en cambio, implicaría que la deriva en una población pequeña durante MIS 4-3 constituye una mejor explicación, también para la expansión de tamaño del cerebro y la adopoción de las tecnologías del Paleolítico Medio entre 300-200 ka.
- En África, los restos de polvo en el Mar Arábigo muestran un largo período de tiempo relativamente húmedo y estable entre 640 y 427 ka. La población debió de alcanzar un gran tamaño que, a través de la selección positiva, habría impulsado la evolución anatómica (el cráneo Bodo, Homo rhodesiensis, que data de 600 ka; Clark et al, 1994; Rightmire 1996) y la innovación cultural (Formación Kapthurin en Lake Baringo; Johnson y McBrearty 2010; Tryon y McBrearty 2006).
- El origen de los humanos modernos se remonta a finales de OIS7-OIS 6, un intervalo hiperárido. La evidencia genética también apoya la hipótesis de que los ancestros de los humanos modernos de África experimentaron un importante cuello de botella poblacional durante este período, aunque algunos datos contradicen la hipótesis. Los primeros fósiles de morfología reconociblemente moderna están datados en 195-160 ka. Se esperaría que la deriva genética fuese el factor dominante durante ese período, pero el clima de África era un mosaico complejo y regionalmente variable (Blome et al. 2012). Estas distintas condiciones podrían explicar la rápida restauración de la variación genética en zonas con cuello de botella poblacional.
- Tanto los datos climáticos como los genómicos sugieren un cuello de botella en el este de África y Arabia después de 60 ka.
Diferencias y semejanzas entre neandertales y sapiens
No hay comentarios:
Publicar un comentario