 |
| OH5 |
El género Paranthropus fue definido en 1938 sobre los fósiles que Broom encontró en Kromdraai en 1936.
Los fósiles asignados al género han sido datados en 2,5-1 Ma.
Grine (1988) y Wood (1991) entre otros, defienden que es un grupo monofilético, mientras que Tobias (1967), Kimbel (1984) y Suwa (1994), entre otros, prefieren clasificarlos dentro de Australopithecus.
Yacimientos
- Kromdraai, Drimolen, Sterkfontein (Sudáfrica): Paranthropus robustus Broom 1938.
- Swartkrans (Sudáfrica): Paranthropus crassidens Broom 1949. Para la mayoría, las diferencias entre los restos de los anteriores yacimientos no justifican un taxón diferente.
- Koobi Fora, Peninj, Chesowanja, Olduvai, Uraha, West Turkana, East Turkana (Kenia): Paranthropus boisei Leakey 1959.
- Omo (Etiopía), West Turkana, East Turkana (Kenia): Paranthropus aetiopicus Arambourg y Coppens, 1967.
| Región | Sitio | Formación geológica | Edad estimada de los homínidos (millones de años) | Método de datación | Los especímenes clave | Especies presentes |
| África del Este | West Turkana, Kenia | Nachukui | 2.5-2.35 | radiométricas, correlación de estratos | KNM-WT 17000 (cráneo), KNM-WT 16005 (mandíbula) | P. aethiopicus |
| 2.3-1.6 | Varios especímenes | P. boisei | ||||
| Koobi Fora, Kenya | Koobi Fora | 2.2-1.88 | radiométricas; tefroestatigrafía; radiometría de rastros de fisión de uranio, correlación de estratos | KNM-ER 1500 (esqueleto parcial) y otros | P. boisei | |
| 1.88-1.65 | KNM-ER 406, 407, 732 (cráneos) y otros | P. boisei | ||||
| 1.65-1.39 | KNM-ER 729, 3230 (mandíbulas) y otros | P. boisei | ||||
| Omo, Etiopía | Shungura | 2.6-2.3 | radiométricas, correlación de estratos | Omo 18-18 (mandíbula edéntula; holotipo de P. aethiopicus ), y otros, la mayoría, dientes aislados | P. aethiopicus | |
| 2.3-1.2 | Varios ejemplares, la mayoría dientes | P. boisei | ||||
| Chesowanja, Kenya | Chemoigut | 2.0-1.5 | Bioestratigrafía; radiométría de la capa de cobertura | CH1 (cráneo parcial), otros fragmentos | P. boisei | |
| Konso, Etiopía | Konso | 1,4 | radiométrica; tefroestatigrafía, correlación de estratos | KGA 10-525 (cráneo), y otros | P. boisei | |
| Malema, Malawi | Chiwondo | 1,5 | bioestratigrafía | RC 911 (maxilar) | P. boisei | |
| Peninj, Tanzania | Humbu | 1.7-1.3 | radiométrica; magnetoestratigrafía | Peninj mandíbula | P. boisei | |
| Garganta de Olduvai, Tanzania | Olduvai | 1.9-1.7 | radiométrica; bioestratigrafía | OH 5 (cráneo; holotipo de P. boisei ) | P. boisei | |
| 1.7-1.2 | Varios especímenes | P. boisei | ||||
| África del Sur | Swartkrans, Sudáfrica | Monte Christo | 1.8-1.5 | bioestratigrafía | > 300 ejemplares aislados, en su mayoría restos dentales, incluyendo SK6 (holotipo de P. crassidens ) | P. robustus( P. crassidens) |
| 1.5-1.0 | P. robustus( P. crassidens) | |||||
| 1.5-1.0 | P. robustus( P. crassidens) | |||||
| Kromdraai, Sudáfrica | Monte Christo | 2.0-1.5 | bioestratigrafía, polaridad invertida | Cerca de 30 especímenes, incluyendo TM1517 (cráneo; holotipo de P. robustus ) | P. robustus | |
| Sterkfontein, Sudáfrica (M5B) | Monte Christo | 1.4-1.1 | magnetoestratigrafía | Dos especímenes: Stw 566 y 569 Stw | P. robustus | |
| Drimolen, Sudáfrica | Monte Christo | 2.0-1.5 | Composición General asociación faunística; no hay fechas absoutas | > 80 homínidos, incluyendo DNH 7 (cráneo casi completo) y DNH 8 (mandíbula masculina) | P. robustus | |
| Gondolin, Sudáfrica | Eccles | 1.9-1.5 | bioestratigrafía (provisional) | GDA-2: muy gran mandibular M2 | P . sp. | |
| Cueva de Cooper, Sudáfrica | Monte Christo | 1.5-1.4 | radiométrica; bioestratigrafía | COB 101 (cráneo parcial) y otros, sobre todo dientes, también postcraneales | P. robustus | |
| Tabla 1: El Paranthropus evidencia fósil. | ||||||
Características
- Bipedación más afirmada que en los australopitecos. Talla algo mayor. Brazos relativamente más cortos y piernas más largas. Anatomía del pie mal conocida.
- Mano similar a la de Homo habilis, con algunas características arcaicas como unas primeras falanges robustas y curvadas, pero el resto de los huesos indican manipulaciones precisas y de presa poderosas.
- Diformismo sexual muy pronunciado, con valores similares a los del género Gorilla.
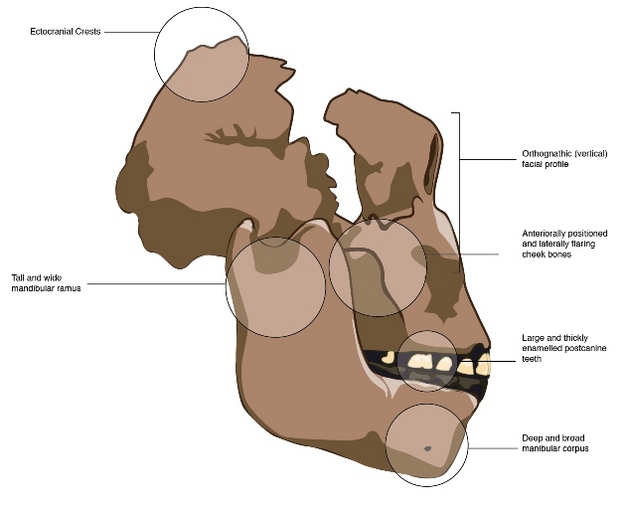
- Enorme rostro cóncavo, muy alto, con mandíbula muy profunda, estructurado para una masticación potente. Los músculos masticadores proyectan hacia delante las partes periféricas del rostro. Huesos malares adelantados, ampliando la superficie de los músculos masetereos. El arco cigomático, en forma de asa redondeada, se curva hacia fuera adquiriendo un mayor diámetro y posibilitando unos músculos temporales de gran tamaño. Cresta sagital para la inserción de poderosos músculos temporales, continuando en una cresta occipital nucal. Los pómulos avanzan hasta ocultar el orificio nasal, que está alojado en una depresión. No hay umbral entre la base nasal y el clivus nasoalveolar. Maxilar en posición más posterior que en el resto de los homínidos, retrasando con ello la zona de los premolares. Mandíbulas muy anchas y gruesas.
- Incisivos y caninos reducidos. Premolares y molares muy grandes y robustos cubiertos de esmalte muy grueso.
- Cerebro desarrollado con 550 cc por término medio, que aumenta un 25% en un millón y medio de años. Lóbulos temporales y frontales más desarrollados que en los primeros Homo. El desarrollo de las áreas parietales empuja el cerebelo hacia delante, al igual que en Homo (homoplasia). Vena meníngea media menos densa que en los australopitecos. El drenaje sanguíneo se efectúa principalmente por la vena occipital.
- P. aethiopicus: 410 cc.
- P. boisei: 400-545 cc.
- P. robustus 533 cc.
Modos de vida
Los parántropos surgen cuando se desarrollan las zonas semiáridas de campo abierto (marchadores) y se extinguen los Australopithecus que son arborícolas. Conviven con herbívoros, carnívoros y otros mamíferos.
Aunque omnívoros como los póngidos y los Australopithecus son fundamentalmente vegetarianos en un 80% de su dieta. El aparato masticatorio, por su potencia, sugiere una dieta vegetariana dura (plantas de hoja y tallo duro, frutos secos), pero salvo en el caso del P.boisei, el análisis de isótopos y de microdesgaste de los dientes hallados no revela una dieta de alimentos duros. Es posible que los parántropos solo recurriesen a los alimentos duros en situaciones de estrés alimenticio o que consumiesen cangrejos y otros mariscos (Martínez et al, 2016; Jordi Marcé-Nogué, 2020).
Balolia, Soligo y Wood (2017), apuntan que la cresta sagital puede haber sido objeto de selección sexual.
De acuerdo con el esqueleto de sus manos pudieron fabricar instrumentos líticos y de acuerdo con hallazgos en niveles estratigráficos similares parece que los construyeron, quizá sólo para machacar huesos. Manejaban palos de cavar.
Balolia, Soligo y Wood (2017), apuntan que la cresta sagital puede haber sido objeto de selección sexual.
De acuerdo con el esqueleto de sus manos pudieron fabricar instrumentos líticos y de acuerdo con hallazgos en niveles estratigráficos similares parece que los construyeron, quizá sólo para machacar huesos. Manejaban palos de cavar.
Taxonomía y filogenia
Rama colateral procedente de la separación (4-5 Ma) desde un antepasado común (¿Ardipithecus ramidus? ¿Orrorin tugenensis?) que se extingue hace 1,5 Ma. Conviven con los Homo habilis durante 1,5 Ma.
Estos son los principales problemas taxonómicos y filogenéticos que afectan al género.
Estos son los principales problemas taxonómicos y filogenéticos que afectan al género.
- ¿Paranthropus es monfilético?
- Para Walker y colaboradores la presencia de rasgos comunes en ejemplares robustos de Sudáfrica y África del Este serían homoplasias y por tanto los australopitecinos robustos formarían dos clados, dos linajes separados:
- La sucesión P. aethiopicus-P. boisei
- P. robustus
- Para Skelton y McHenry, hay relación filogenética entre P.boisei y P. robustus, con exclusión de P. aethiopicus.
- Esta teoría tiene pendiente explicar cómo el menos derivado (es decir, menos robusto P. robustus ha evolucionado a partir del más antiguo y derivado P. boisei.
- Estudios a favor de la monofilia:
- Wood, 1988.
- Corruccini, 1994.
- Strait et al, 1997.
- Strait y Grine, 2004.
- Kimbel et al, 2004.
- Villmoare et al, 2014.
- Razones para reexaminar la monofilia de Paranthropus.
- El descubrimiento de KNM-WT 17000 y otros especímenes, con rasgos en mosaico de Australopithecus afarensis y Paranthropus.
- Los hallazgos se concentran en unas pocas regiones del cuerpo y puede que no reflejen las verdaderas relaciones evolucionarias.
- La mayor parte de las características que relacionan a los parántropos entre sí, se relacionan con la masticación y son interdependientes. Podrían ser resultado de una homoplasia, un proceso de evolución convergente. Por ejemplo, la megadontia en una cara relativamente pequeña se da también en Australopithecus garhi.
- Se han realizado estudios para probar la monofilia de Paranthropus, sin resultados claros:
- Wood, 1988.
- Turner y Wood, 1993.
- ¿P. robustus y P. boisei pueden agruparse en la misma especie?
- El hallazgo en 1997 en Konso (Etiopía) del cráneo KGA 10-525 hace pensar que rasgos considerados fundamentales pueden ser en realidad polimorfismos. El P. boisei de Olduvai sería un espécimen extremo mientras los descubiertos en Turkana y Konso tendrían una morfología intermedia entre los de Olduvai y los P. robustus sudafricanos. Además, descubrimientos recientes en el sur de África (Drimolen y Gondolin) parecen llenar la separación entre robustus y boisei.
- Sin embargo, Wood y Lieberman (2001) y Constantino y Wood (2004) han confirmado las diferencias entre los hallazgos del Este de África y los del Sur.
- Wood y Constantino consideran que las similitudes con Australopithecus africanus son un caso notable de homoplasia con ambos linajes tendiendo hacia la hipertrofia morfológica masticatoria y megadoncia extrema de los postcaninos, y además en ausencia de similitudes en la dieta.
- ¿P. aethiopicus y P. boisei pueden agruparse en la misma especie?
- Dos estudios han analizado este problema en detalle:
- Suwa (1988) encontró una autapomorfia en P. boisei: coronas de los premolares mandibulares con talónido particularmente largo y elaborado.
- Wood et al (1994) localizaron más diferencias en la mandíbula y en la dentición mandibular, como la altura de la sínfisis mandibular, el corpus mandibular en las áreas de las coronas de M1 y P4.
- P. aethiopicus es más prognato, con los incisivos más largos, la base craneal menos flexionada y los postcaninos son más pequeños.
- La naturaleza de las diferencias es consistente con el concepto de cronoespecie.
- ¿Hay relación filogenética con Australopithecus afarensis?
- Kimbel et al (2004) y Rak y Kimbel (2006) no encontraron evidencia de relaciones filéticas directas.
- ¿Hay relación filogenética con Australopithecus africanus?
- No es posible porque africanus se extiende hasta hace 2,3 Ma mientras que aethiopicus aparece hace 2,6 Ma.
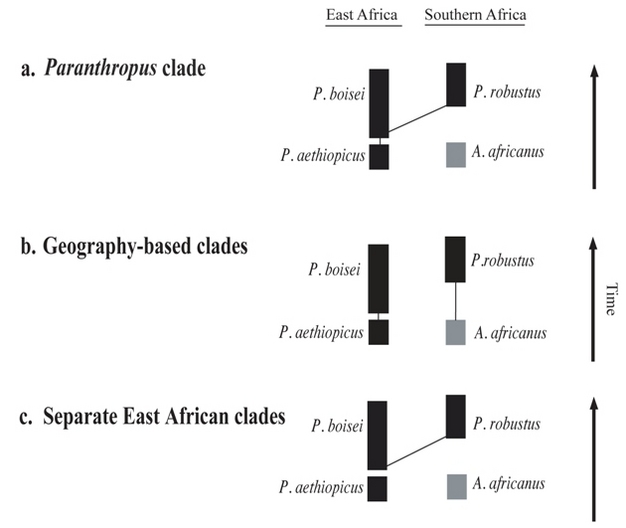 |
| (a) los "robustos" homínidos de África oriental y meridional forman un clado monofilético y por lo tanto pueden ser agrupadas en el género Paranthropus (Grine) (b) Los homínidos "robustos" se originó de forma independiente en el este y sur de África (Walker) (c), propuesto originalmente por Skelton y McHenry. Hay relación filogenética entre P.boisei y P. robustus, con exclusión de P. aethiopicus . |
- Se habían vuelto demasiado especializados para hacer frente a las cambiantes condiciones ambientales.
- La competencia con los Homo.
Hay pocos indicios a favor de cualquiera de ellas.
Te puede interesar:
|
No hay comentarios:
Publicar un comentario