 |
| Localización de los Hominini más antiguos (incertae sedis en el caso de Sahelanthropus). © 2013 Nature Education. imágenes adaptadas de Brunet et al. 2002 (Sahelanthropus), Semaw et al. 2005 (Ardipithecus ramidus Gona), Haile-Selassie 2001 (Ardipithecus kadabba), White et al. 2009 (Ardipithecus ramidus), Senut et al. 2001 (Orrorin). |
Orrorin tugenensis (Senut et al, 2001)
- Holotipo: BAR 1000 '00, mandíbula en dos fragmentos:
- BAR 1000a 00: fragmento del lado izquierdo de mandibula, con M2–3.
- BAR 1000b00: fragmento del lado derecho de mandíbula, con M3.
- Yacimientos: Cuatro localidades en la Formación de Lukeino, distrito de Baringo, Kenia (Cheboit, Kapsomin, Kapcheberek y Aragai).
Orrorin significa “primer hombre” en el idioma de Tugen.
Además del holotipo, los hallazgos incluyen:
- Un fragmento de la parte proximal del fémur (BAR 1002’00), que comprende 2/3 del hueso completo.
- Fragmentos de otros dos fémures.
- Fragmento distal de radio.
- Fragmento proximal de la falange de un dedo.
- Tres fragmentos de una mandíbula.
- Dientes aislados.
Estos restos, custodiados tras su descubrimiento en una instalación local, han desaparecido.
 |
| Orrorin tugenensis |
 |
| Fémures de ( A ) P. troglodytes ,( B ) Orrorin (BAR 1002'00), ( C y D ) Paranthropus robustus (SK 97 y SK 82, invertido), ( E ) A. afarensis (AL 288-1AP), ( F ) Paranthropus boisei (KNM-ER 1503, invertido), ( G ) Homo temprano (KNM-ER 1481), y ( H ) H. sapiens moderno. Los homínidos tempranos (B a F) se diferencia de los grandes simios y de los humanos modernos en que el cuello del fémur es largo y estrecho. Los Homo tempranos (G) tienen fémures con grandes cabezas y cuellos amplios. Barra de escala, 2 cm. Science. |
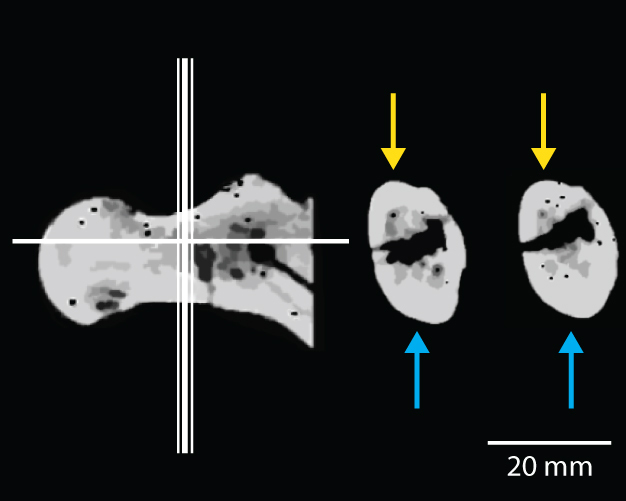 |
| CT-scan del cuello femoral de BAR 1002'00, Orrorin tugenensis. A la izquierda se muestra dónde se tomaron las secciones. La distribución del hueso cortical es asimétrica: la corteza es mucho más gruesas inferiormente (flechas azules) que superiormente (flechas amarillas). Los seres humanos también tienen una distribución asimétrica similar del hueso cortical, mientras que los monos africanos tienen una distribución mucho más uniforme debido a las diferentes tensiones de carga de bipedación y el cuadrupedalismo. © 2013 Nature Education Adaptación de Pickford et al, 2002. |
 |
| BAR 1002'00 Orrorin tugenensis. David Alba. |
Adam van Arsdale: Orrorin tugenensis
Fragmentos de Chemeron.
Fragmentos hallados en Chemeron, lago Baringo, Kenia. Datados en 5,1 Ma.- Extremo proximal de húmero (1980).
- La Mandíbula de Tamarin (1984) es un fragmento mandibular datado en 4,42-4,3 Ma (KNM-TH 13150: Hill 1985; Deino et al. 2002) hallado en Tugen Hills (Kenya). En algunos autores aparece asignada a Ardipithecus ramidus (Kissel y Hawks, 2015). La inclusión de la Mandíbula de Tabarin en Ardipithecus tendría como consecuencia una extensión geográfica sustancial del género.
Fragmento de mandíbula de Lothagam.
El fragmento de mandíbula de Lothagam, Kenia (KNM-LT 329), en el que se conserva el alveolo del P4, el M1 y la raíz distal de M3, fue descubierto en 1967 y publicado por Patterson en 1970. Su edad se estima entre hace 5,2-4,2 Ma. Una edad más antigua de 4,4 Ma la colocaría fuera del rango conocido para Austrolopithecus anamensis.Según Kissel y Hawks (2015), la mandíbula de Lothagam, no corresponde a Ardipithecus. Sus dimensiones molares, están claramente fuera del rango conocido para ese género, pero quedan dentro del rango de Australopithecus afarensis y Australopithecus anamensis. Si se confirmara una datación del espécimen más a antigua que hace 4,4 Ma, estaríamos ante una evidencia de que Australopithecus coexistió con Ardipithecus y posiblemente con otros Hominini.
 |
Marco superior izquierda: Comparación del canino superior y premolar de Ar. kadabba con los de un chimpancé hembra. Nótese la forma grande y triangular del canino superior.
Marco superior derecha: Comparación del canino superior del Ar. kadabba con los de Ar. ramidus y Au. afarensis.
Marco inferior: Comparación de la falange proximal del pie de Ar. kadabba con las del chimpancé y Au. afarensis. Las flechas señalan la presencia de una inclinación dorsal en Ar. kadabba y Au. afarensis que se observa en los seres humanos modernos, pero no en monos.
© 2013 Nature Education. Marco superior adaptado de Haile-Selassie et al, 2004; Marco inferior adaptado de Haile-Selassie et al, 2009.
|
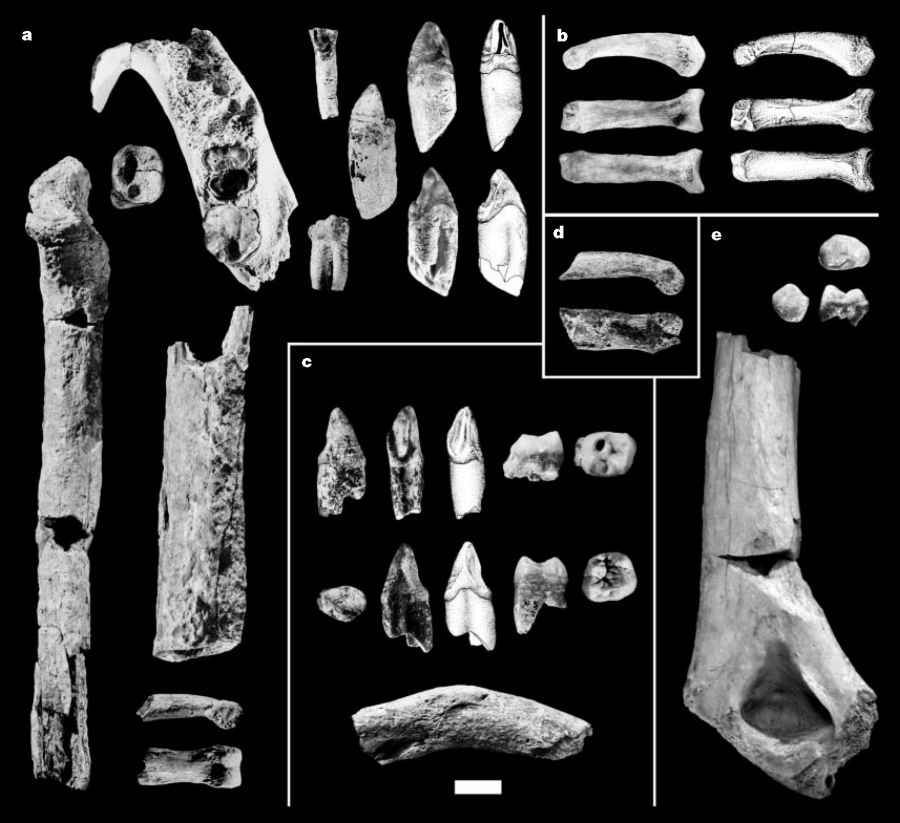
A. ARA-VP-2/10, mandíbula y dientes asociados; ARA-VP-2/120, cúbito y eje del húmero; ARA-VP-2/11, falange de la mano.
B. AME-VP-1/71, vistas lateral, inferior y dorsal de la falange del pie.
C. STD-VP-2, dientes y clavícula parcial.
D. DID-VP-1/80, falange de la mano.
E. ASK-VP-3/160, vista oclusial, mesial y bucal; ASK-VP-3/78, vista posterior.
(Haile-Selassie, 2001).
A. ARA-VP-2/10, mandíbula y dientes asociados; ARA-VP-2/120, cúbito y eje del húmero; ARA-VP-2/11, falange de la mano.
B. AME-VP-1/71, vistas lateral, inferior y dorsal de la falange del pie.
C. STD-VP-2, dientes y clavícula parcial.
D. DID-VP-1/80, falange de la mano.
E. ASK-VP-3/160, vista oclusial, mesial y bucal; ASK-VP-3/78, vista posterior.
(Haile-Selassie, 2001).
Ardipithecus kaddaba (Haile-Selassie et al 2004).
- Holotipo: ARA-VP-2/10.
- Yacimientos: Localidades de la orilla occidental del curso medio del río Awash (Saitune Dora, Alayla, Asa Koma y Digiba Dora).
En 2002 se descubrieron seis dientes más, datados en 5,77-5,54 Ma. Otros dientes, datados en 6,3 Ma, no han podido ser asignados a ninguna especie (Scott W. Simpson et al, 2015)
Si bien en principio estos restos fueron asignados a A. ramidus, el hecho de que entre el canino superior y el P3 inferior se diese una relación oclusal única entre los Hominini, justificó la definición de una nueva especie.
 |
| A. En el cráneo, los chimpancés combinan una parte posterior ancha primitiva con un alargamiento de la parte anterior de la base del cráneo derivado. Los homínidos comparten una base del cráneo acortada derivada. B. En la dentición, los homínidos perdieron las características del conjunto C/P3 basadas en un canino sobresaliente en los machos. C. En las extremidades, Ardipithecus ramidus es primitivo. El chimpancé es derivado en su brazo alargado y el humano en la pierna alargada. D. En la pelvis, los homínidos comparten unas palas ilíacas inferiores más amplias y una posición baja de la articulación del sacro. E. La mano del chimpancé muestra unos metacarpianos alargados y reforzados para la escalada y la suspensión. Los homínidos, unos metacarpianos cortos, primitivos, y unas falanges acortadas. F. El pie de Ardipithecus ramidus muestra una parte media primitiva. Los simios, una estructura flexible acortada. Los humanos, una estructura de palanca reforzada que absorbe al mismo tiempo los impactos. |
Ardipithecus ramidus (White, Suwa y Asfaw, 1994-1995).
- Holotipo: ARA-VP-6/1.
- Yacimientos:
- Aramis, al oeste del curso medio de río Awash (Afar, Etiopía). Localidades 1, 6 y 7.
- As Duma, Gona Western Margin (GWM), Afar, Etiopía.
- Localidad GWM3, Abdu Mohamed Ali
En enero de 2005 se informó del descubrimiento de los restos de por lo menos nueve individuos clasificados como Ardipithecus ramidus, de entre 4,4 y 4,1 millones de años de antigüedad, en As Duma, al norte de Etiopía, por el equipo de la Universidad de Indiana dirigido por Sileshi Seaslug (Semaw et al, 2005).
En 2009 se informó del hallazgo de nuevos restos en Aramis, incluyendo ARA-VP-6/500, Kuseralee Dora y Sagantole.
En la lengua afar, local, ardi significa “tierra, suelo” y ramis “raíz”.
- ARA-VP-6/1 Diez molares de un individuo.
- ARA-VP-7/2 Tres huesos enteros del brazo izquierdo correspondientes a un individuo, con rasgos en mosaico.
- ARA-VP-6/500. Esqueleto fósil relativamente completo hallado en 1994 en Aramis. Corresponde a una hembra (Ardi). Datado hace unos 4,4 Ma (3,9 según Beagle y Kappelman).
- GWM3/P1. Fragmento de mandíbula recuperado en 1999.
- GWM3/P2. Fragmento proximal de falange de la mano. Simpson et al, 2019.
- GWM5sw/P56. Mandíbula.
- GWM5sw/P58. Fragmento intermedio de falange de la mano. Simpson et al, 2019.
- GWM5sw/P701. Escafoides mano izquierda. Simpson et al, 2019.
- GWM10/P1. Falange manual casi completa. Semaw et al., 2005. Simpson et al, 2019.
- GWM67/P1. Fragmento de maxilar recuperado en 2005.
- GWM67/P2. Esqueleto parcial (que incluye fragmentos de ambos miembros inferiores, un pequeño fragmento de la espina vertebral y posiblemente algunos fragmentos de costilla), datado en 4,8-4,3 Ma y recuperados entre 2006-2013. El tobillo y hállux muestran una mayor adaptación a la bipedación que los restos estudiados anteriormente (Simpson et al, 2019).
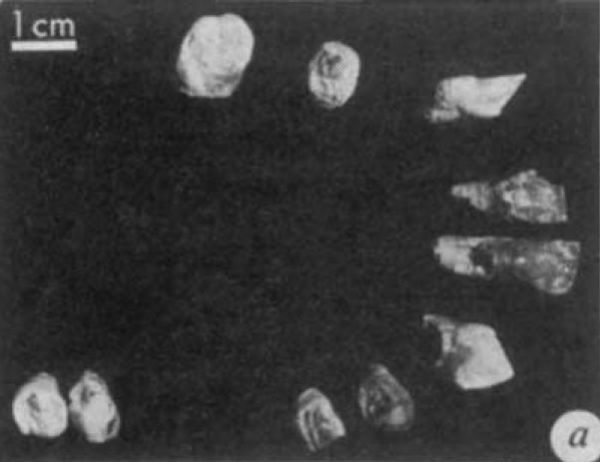 |
| ARA-VP-6/1, dentición superior e inferior de un mismo individuo (White, et al. 1994) |
 |
| Izquierda, Sahelanthropus (Zollikofer et al. 2005) Derecha, Ardipithecus ramidus (Suwa et al. 2009). |
 |
| Ardi y reconstrucción artística de J. H. Matternes. 2009. |
Adam van Arsdale: Ardipithecus
Enlaces:
- Gómez Castanedo, A: (2012), "Los orígenes del género Homo. Una exploración de la innovación como motor evolutivo". Revista Nivel Cero, 13. Santander.
- The Earliest Hominins: Sahelanthropus, Orrorin, and Ardipithecus By: Denise F. Su (Curator and Head, Department of Paleobotany and Paleoecology, Cleveland Museum of Natural History) © 2013 Nature Education
- Ardipithecus ramidus and the Paleobiology of Early Hominids. Tim D. White, Berhane Asfaw, Yonas Beyene, Yohannes Haile-Selassie, C. Owen Lovejoy, Gen Suwa, Giday WoldeGabriel
- Reexamining Human Origins in Light of Ardipithecus ramidus. C. Owen Lovejoy
Te puede interesar:
|