como instrumento de interpretación de la evolución humana, afectada en gran medida por los cambios climáticos globales.
Durante mucho tiempo, el
gradualismo filético fue el modelo evolutivo privilegiado en la Paleoantropología, dando lugar a un patrón filogenético lineal, donde era imposible la coexistencia de varias especies en el mismo tiempo y lugar (Dobzhansky, 1962; Mayr, 1963). Actualmente, el crecimiento de la base empírica, junto con evidencias convergentes de la Paleontología, la Biología Molecular, la Paleobiogeografía, la Paleoecología y otros campos, rechazan el modelo lineal.
Los datos paleontológicos disponibles actualmente, apuntan a la presencia entre 7-4,4 Ma de al menos cuatro especies candidatas a Hominini temprano:
Sahelanthropus tchadensis, Orrorin tugenensis,
Ardipithecus kaddaba y
Ardipithecus ramidus. Aunque el consenso no es total, la mayoría está de acuerdo en incluirlas en el clado Hominini en base a una serie de rasgos derivados compartidos que sugieren modificaciones conductuales significativas: reducción de tamaño y otros cambios en la morfología de los caninos, insinúan cambios en la dieta; la localización y orientación del
foramen magnum y varios huesos postcranelaes, en la postura y la forma de desplazamiento. Al mismo tiempo, muchos rasgos de los pies, extremidades superiores, pelvis, capacidad craneal, etc, no muestran innovación o muy poca. Esta mezcla de rasgos derivados y retenciones parece estar relacionada con la experimentación de formas híbridas de locomoción, quizá alternado la bipedación con la cuadripedia o quizá una bipedación por encima de las ramas de los árboles, en el momento de la separación con el linaje de los chimpancés. El área biogeográfica donde aparecen estas especies cubre desde el Rift hasta el Sur de África, si bien
tchadensis fue encontrado 2.500 km al oeste del Valle del Rift. Los datos recogidos de los carbonatos del suelo, isótopos de carbono
n-alcanos en sedimentos marinos, dientes fosilizados de mamíferos y simulaciones matemáticas, establecen una tendencia a la aridez a largo plazo en el este de África, modulada por periodos de alta variabilidad climática y cortos periodos de relativa estabilidad. La aridez fue debida a un calentamiento global que comenzó al final del
Paleoceno y la variabilidad climática a la acción intermitente de los ciclos de Milankovitch y la progresiva separación y levantamiento tectónico del Este de África, que fue taponando las corrientes de aire seco y caliente procedentes del Océano Atlántico. Estos cambios climáticos y ambientales transformaron el este y el sur de Africa en una región heterogénea con altas montañas y un mosaico de hábitats desde bosques húmedos y cerrados a praderas y desiertos. Las nuevas adaptaciones de los Hominini tempranos parecen asociadas directamente con un contexto inestable y fluctuante y presiones selectivas relacionadas con el incremento de la extensión de los hábitats abiertos.
El género
Australopithecus aparece hace 4,2-4 Ma, con similares condiciones ambientales.
Australopithecus anamensis estaba más adaptado que
Ardipithecus a hábitats más áridos. El género
Australopithecus comprende un número significativo de especies que poblaron el este y el sur de África, aunque varias son controvertidas, y se extendió por un largo periodo entre 4,2-1,8 Ma. Cada una de estas especies muestra una mezcla única de características anatómicas incluyendo retenciones y algunas innovaciones, sobre todo en la dentición y la postura, que anticipan al género Homo. Estos rasgos en mosaico indican una dieta diversificada y una bipedación avanzada en el marco de una mezcla de modos de locomoción, con adaptaciones a la trepa y a la vida en los árboles, una flexibilidad para hacer frente a un entorno cambiante con inestabilidad persistente del clima. La industria Lomekwiense se ha datado en hace 3,3 Ma, conjeturalmente en asociación con
Kenyanthropus platyops.
Cuando surgió el género Homo, la bipedación se convirtió en prevalente y luego obligada. Esta transición evolutiva pudo llevarse a cabo, con un patrón en mosaico, por unas pocas especies morfológicamente inestables o por una pluralidad de especies separadas, cada una con un conjunto específico de características. La inestabilidad climática y la fragmentación del hábitat fueron los retos macroevolutivos. Una pluralidad de estrategias de adaptación, compartiendo la misma flexibilidad, fue la respuesta. En términos de una teoría de múltiples niveles evolutivos, vemos un conjunto de comportamientos experimentales, desencadenados por fuerzas externas que operan a nivel macro-evolutivo: las fluctuaciones en el clima global y los cambios ambientales regionales. A nivel inferior, cada especie buscó su propio camino y estrategias de flexibilidad ecológica y especialización dietética, produciendo diferentes conjuntos de cambios adaptativos.
En los estallidos de diversificación de hace 2,9-2,6 Ma y 2,4-2 Ma, la ramificación de los Hominini alcanzó su mayor extensión, desmintiendo cualquier teoría lineal para la filogenia humana. Tras una aparente estasis de un millón de años, y aunque otros miembros del género le sobreviven durante otro millón de años,
Australopithecus afarensis se extinguió hace 2,9 Ma y el género
Homo parece surgir poco después: la mandíbula de Ledi-Geraru, ha sido datada en 2,8-2,75 Ma. El género
Paranthropus aparece hace 2,7 Ma. Observamos, por tanto, una sorprendente cohabitación de diferentes géneros en el este y el sur de África a comienzos del Pleistoceno. Estas poblaciones muestran un nuevo mosaico de rasgos relacionados con los nichos ambientales. La impresionante radiación de especies junto con la inestabilidad climática y la fragmentación del hábitat sugieren fuertemente una especiación geográfica y un cambio puntuado.
A comienzos del Pleistoceno se produjo un enfriamiento global relacionado con la variación de la órbita de la Tierra y la inclinación del eje de giro, asi como por la deriva de los continentes que dio lugar al istmo de Panamá o, en una hipótesis alternativa, al cierre de la vía marítima por Indonesia. Las corrientes se modificaron, enfrió la superficie del mar, se desviaron los vientos y los hielos polares se expandieron, provocando una severa Edad del Hielo, con intensas fluctuaciones entre fases glaciares e interglaciares. Debido a estos cambios globales, el clima de África se hizo todavía más seco e inestable, alternando ciclos de alta y baja variabilidad climática de humedad-aridez dando lugar a periodos de relativa estabilidad rotos por intervalos de hábitats impredecibles y recursos inciertos, con una coexistencia de nichos ecológicos heterogéneos. El Sahara se extendió y aumentó la proporción de plantas C4. A causa de esta fragmentación ambiental, muchas poblaciones periféricas quedaron aisladas, lo que produjo las condiciones ideales para la extinción de especies, especiaciones alopátridas, radiaciones adaptatativas y evolución puntuada.
- Paranthropus estaba equipado con un potente aparato masticatorio para una dieta vegetariana casi exclusiva en plantas C4. P. boisei, más especializado en los pastos y juncias y P. robustus en las frutas carnosas de árboles. Esta ampliación de la gama alimenticia permitió la subsistencia en ambientes en mosaico.
- En el mismo período y en el mismo entorno inestable, Homo desarrolló estrategias completamente diferentes, que revelan una mayor flexibilidad de comportamiento, con cerebros más grandes, reducción del tamaño de los dientes posteriores, flexibilidad en la dieta y producción de artefactos de piedra.
- La Hipótesis Turnover-Pulse propone que las especies aparecen y desaparecen cuando sus entornos físicos se ven sometidos a cambios muy significativos. Según Elisabeth Vbra (1980), los ecosistemas experimentan periódicamente interrupciones importantes provocadas por cambios ambientales físicos que generan extinciones en masa. Las extinciones, a su vez, facilitan nuevas oportunidades a las especies más generalistas que pueden migrar a otros entornos, produciéndose una especiación al azar en áreas aisladas (alopátrida). Por lo tanto, la rotación de la mayoría de linajes se produjo en forma de pulsos, casi sincrónicos entre los diversos grupos de organismos. Los cambios pueden ser generalizados, pero en su mayoría son locales y aleatorios en el tiempo.
- La Hipótesis de Potts de la Selección de Adaptaciones a la Variabilidad (1996, 1998, 2013, 2015; Grove, 2011a, 2011b) aboga por el papel de la imprevisibilidad del medio ambiente en la selección de un comportamiento flexible. Desarrolla la hipótesis Turnover-Pulse. Enfatiza la importancia de los sistemas fluctuantes para la introducción de pluralidad genética, selección natural, respuesta adaptativa e innovaciones faunísticas. Esta hipótesis difiere sustancialmente de aquellas que defienden que la evolución está relacionada con un tipo específico de hábitat. Potts propone la existencia de unas adaptaciones a la variabilidad consistentes en estructuras y comportamientos que proporcionan la necesaria flexibilidad para dar respuesta a cambios complejos en el entorno y que son uniformes dentro de las especies, pero que son capaces de mediar rasgos fenotípicos secundarios variables. Estas adaptaciones surgen en poblaciones aisladas y la selección organísmica las promueve o al menos las conserva. Cuando se producen cambios climáticos bruscos, los organismos con estas adaptaciones sobreviven a los extremos climáticos. La variabilidad climática en ciclos largos se convierte así en un agente de selección de adaptaciones a la variabilidad.
Australopithecus y Paranthropus se extinguieron después de hace 2 Ma, pero Homo sobrevivió y dio lugar a una radiación adaptativa a escala global. Homo ergaster presenta muchos rasgos innovadores que incluyen la expansión de tamaño del cerebro, el acortamiento del intervalo entre nacimientos, el retraso en el desarrollo, el tamaño y forma corporal, la anatomía postcraneal y la flexibilidad en la dieta. Todas estas innovaciones parecen estar relacionadas con una adaptación versátil y la capacidad de colonizar nuevos ambientes, incluyendo el Out of Africa.
- Se ha propuesto una interesante hipótesis ecológica para explicar esta dispersión. En las fases húmedas de las fluctuaciones climáticas, el Sahara atrajo a los grandes herbívoros y a sus depredadores, incluyendo Homo. Cuando las cuencas de los lagos se secaron, los Homo se vieron obligados a moverse hacia el norte y el sur y algunas áreas refugio pudieron facilitar la especiación alopátrida, inluyendo Eurasia.
Lordkipanidze et al (2013) han propuesto la existencia de una única especie polimórfica hace ca 2 Ma (H. erectus ergaster georgicus), pero esta hipótesis es menos sostenible que la que argumenta la existencia de tres o más especies diferentes: los Homo tempranos experimentaron diferentes morfologías para los distintos contextos ambientales.
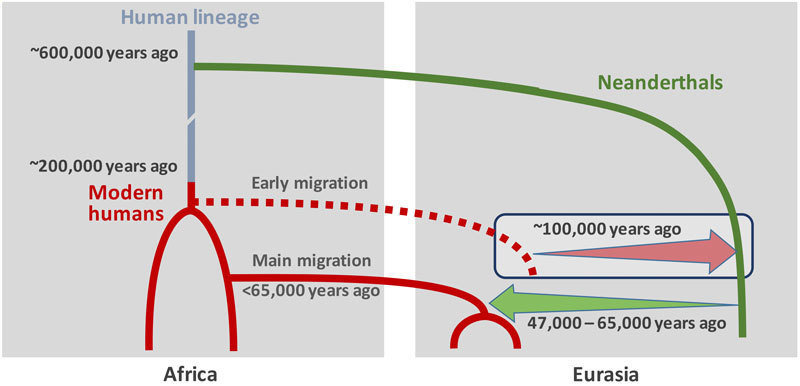
También fue influenciada la especiación de Homo sapiens en África hace ca 200 ka por la inestabilidad climática y las fases áridas. El nacimiento de la cognición moderna, sin embargo, parece más reciente, como si la anatomía y el comportamiento se hubiesen desajustado temporalmente. Solo alrededor de hace 80-60 ka aparecen en África indicios de expresiones simbólicas, quizá asociados a un crecimiento de la población. En Europa aparecen comportamientos simbólicos mucho más tarde, hace 40 ka, en asociación con los cromañones y también en Sulawesi en la misma fecha, lo que es indicio de unos orígenes comunes profundos más que de una convergencia cultural. El bajo nivel de diversidad genética observada en los humanos actuales no africanos puede estar relacionado con cuellos de botella poblacionales durante las crisis climáticas y las olas de expansión Out of Africa hace 120-100 Ka, 85-50 ka y 60-50 ka. Únicamente esta tercera ola final dio lugar a efectos fundadores y difusión planetaria. Toda la población humana no africana desciende e un mismo grupo fundador africano. La aparición de comportamientos simbólicos coincide significativamente con esta oleada: las tecnologías Still Bay y Howieson's Poort confirman la ocurrencia de ráfagas efímeras y puntuadas de innovación tecnológica y de comportamiento, relacionadas con las fluctuaciones climáticas y demográficas: la expansión y contracción de las poblaciones afectó a las redes sociales y a la innovación cultural. Es posible que la inteligencia simbólica fuese facilitada por un lenguaje completamente articulado surgido en el Sur de África en este periodo crucial. Una chispa de ingenio humano (Jacobs y Roberts, 2009) pudo haber conferido a la correspondiente población una ventaja competitiva en términos de tecnología, eficiencia en la comunicación, cohesión social y coordinación grupal, encendiendo y alimentando las conductas innovadoras y la expansión geográfica. Otras poblaciones y especies pudieron haber seguido diferentes trayectorias simbólicas.