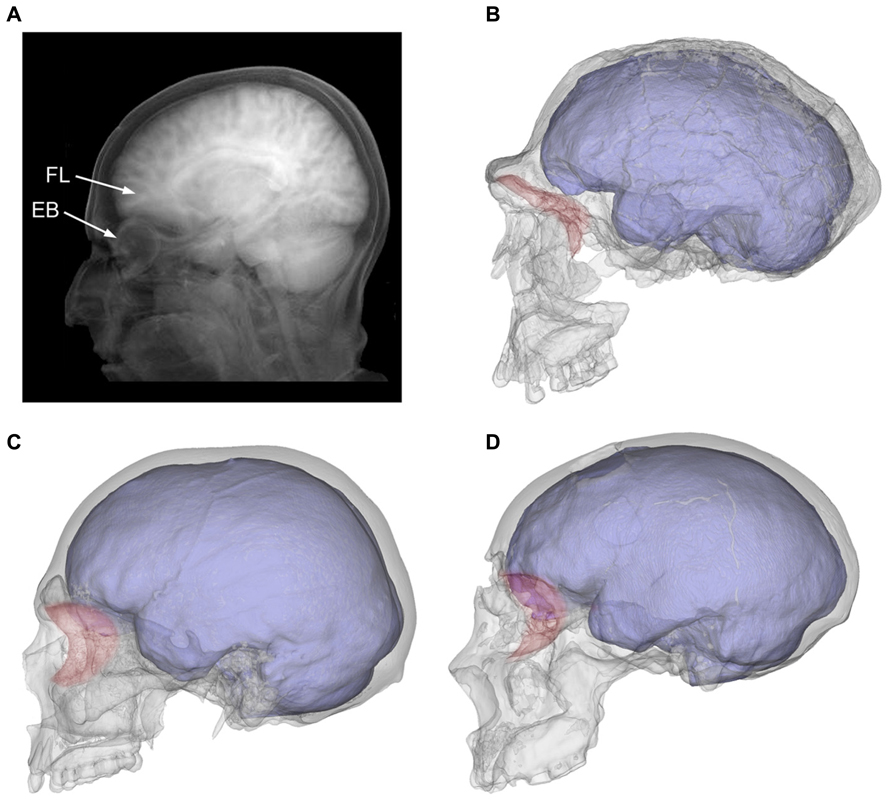
 |
A). La proyección tomográfica muestra la proximidad íntima entre los lóbulos frontales (FL) y el globo ocular (EB) en un individuo humano moderno.
B). Las órbitas (en rojo) se separaron de la fosa craneal anterior en los humanos arcaicos, como Homo ergaster (KNM-ER 3733)
Las órbitas se situaron bajo las áreas prefrontales en los HAM (C; europea moderna) y los neandertales (D; Saccopastore 1).
También se puede apreciar cómo los lóbulos temporales ampliados se asocian más estrechamente con la órbita posterior en los humanos modernos en comparación con estos otros grupos taxonómicos.
|
A pesar de más de cien años de investigación todavía no se sabe qué causa el astigmatismo y la miopía, y por qué se encuentran en correlación con factores tales como el sexo, ascendencia, edad, inteligencia y estatus socioeconómico
Los seres humanos hemos experimentado una historia morfológica única entre los mamíferos, con millones de años de expansión cerebral y reducción del prognatismo facial que han provocado que los ojos y las órbitas descansen directamente debajo de la corteza frontal. Ciertas formas de reducción de la agudeza visual pueden estar relacionadas con la competencia entre los tejidos blandos de los ojos y los duros de los huesos de las órbitas.
El aumento absoluto y relativo del tamaño del cerebro conlleva la modificación de las estructuras craneoespinales y faciales subyacentes, y se asocia con una cara más corta en sentido anteroposterior. La medida en que la totalidad del cráneo ha girado y la cara y las órbitas se han escondido bajo el cerebro, es una característica derivada única de los humanos anatómicamente modernos.
El sulcus y los patrones girales en los lóbulos frontales que se pueden observar en los moldes endocraneales de los homínidos desde hace 2 Ma, revelan cierta evidencia de menor importancia de cambios paleoneurológicos. En los neandertales y HAM se manifiesta un marcado incremento en las proporciones generales, con áreas prefrontales relativamente más anchas en la tapa de Broca, en comparación con otras especies humanas. Es probable que la redistribución lateral de esta masa cortical esté relacionada con las restricciones entre cerebro y estructuras craneales, y, en particular, restricciones impuestas por la arquitectura facial. Los neandertales, con una cara más grande, disfrutaban de áreas frontales más amplias que las de los HAM, aunque las diferencias no son significativas, lo que puede ser debido al tamaño limitado de la muestra.
Por lo tanto, en términos evolutivos, la fosa anterior es libre de cambiar lateralmente, ya que en el género humano las órbitas se desplazan hacia delante, y el músculo temporal es extremadamente pequeño debido a una marcada reducción de tamaño de la boca u hocico. Por el contrario, el aumento vertical de las áreas frontales se ve comprimido en estos dos taxones por una limitación biomecánica específica, y es que en los humanos modernos y los neandertales los lóbulos frontales se encuentran directamente en la parte superior del techo de la órbita. La interacción entre estos componentes anatómicos ha comenzado en los dos linajes de forma independiente hace 150-100 ka.
La encefalización de los homínidos se ha traducido en una serie de modificaciones en la bóveda craneal, base craneal, y cara. Una posición más anterior y el desarrollo lateral de la corteza prefrontal sobre los ojos, la expansión de los lóbulos temporales y la reducción del prognatismo facial, pueden perjudicar la agudeza visual, ya que limitan el desarrollo orbital y ocular. Las restricciones sobre las órbitas y los tejidos blandos de la vista, asociada con la expansión y reubicación anterior de los lóbulos frontal y temporal, podría haber sido más severa en los humanos modernos y los neandertales que en los homínidos pequeño cerebro, como resultado de la relación espacial única entre estos rasgos anatómicos en estos grupos.
Estos cambios evolutivos, además de una disminución comprobada en el volumen orbital desde el Neolítico en China, son las tendencias más importantes a tener en cuenta en la investigación de la mayor frecuencia y gravedad de ciertas formas de reducción de la agudeza visual como la miopía y el astigmatismo.
Curiosamente, disminución de la agudeza visual también es común entre las numerosas razas de perros como el Caniche Toy, Schnauzer Miniatura, Pug, Rottweiler, Collie y Labrador Retriever. Las variedades con una mayor frecuencia y mayor severidad de los defectos de refracción también tienden a ser las que han sufrido mayores fuerzas selectivas humanas en la arquitectura craneofacial, por razones funcionales y/o estéticas: se caracterizan generalmente por pedomorfismo craneofacial y poseen un neurocraneo más frontal y globular, la cara más corta y los ojos prominentes.
Mientras que el crecimiento del globo ocular va paralelo con el crecimiento del cerebro y ambos se cree que son el producto de un mismo gen pleiotrópico, la órbita crece con el resto del cráneo. Por tanto, existe una relación alométrica negativa entre el ojo y la órbita. La reducción bien documentada en tamaño humano en general y robustez que comenzó hace 12 ka actuaría para aumentar el porcentaje de la órbita ocupada por el ojo, hasta el punto de tener que competir por el espacio. La disminución observada en el volumen orbital en Asia oriental podría exacerbar una tendencia existente hacia un mayor tamaño relativo del ojo dentro de la órbita, y potencialmente ayudar a explicar la inusual alta frecuencia de la miopía en esta región geográfica.
La enfermedad de Alzheimer se caracteriza por la acumulación de placas neuríticas compuestas por depósitos extracelulares de péptido amiloide, lo que conduce a la pérdida neuronal, atrofia cortical, y el deterioro cognitivo. A pesar de que las placas son consideradas como un factor causal fundamental de la enfermedad, la cantidad de placa no se correlaciona con alteraciones cognitivas. Los ovillos neurofibrilares, con mayor frecuencia en las áreas temporales mediales, son otra característica principal neuropatológica. Debido a que la atrofia en las primeras etapas de la enfermedad, es mayor en las áreas temporales mediales, los estudios se han centrado en los lóbulos temporales. Más recientemente, sin embargo, se ha demostrado que en las primeras etapas de la enfermedad, se pueden encontrar en la corteza parietal medial alteraciones metabólicas, neuroanatómicas, y cambios histológicos.
La enfermedad de Alzheimer se sufre sólo nuestra especie. Aunque la vida útil extendida en los seres humanos puede implicar problemas de mantenimiento de la integridad del cerebro, la especificidad y la alta prevalencia en nuestra especie pueden sugerir un vínculo evolutivo entre su patología y nuestra biología cerebral. En cualquier caso la neurodegeneración en la edad avanzada no afecta la aptitud genética, ya que influye en un período que generalmente va más allá de las etapas de la reproducción humana. En términos de evolución, esta enfermedad puede ser interpretada como un caso de pleiotropía antagónica, en la que las ventajas cognitivas son pagadas con los costos de un sistema anatómico de gran alcance, caro y delicado.
El sulcus y los patrones girales en los lóbulos frontales que se pueden observar en los moldes endocraneales de los homínidos desde hace 2 Ma, revelan cierta evidencia de menor importancia de cambios paleoneurológicos. En los neandertales y HAM se manifiesta un marcado incremento en las proporciones generales, con áreas prefrontales relativamente más anchas en la tapa de Broca, en comparación con otras especies humanas. Es probable que la redistribución lateral de esta masa cortical esté relacionada con las restricciones entre cerebro y estructuras craneales, y, en particular, restricciones impuestas por la arquitectura facial. Los neandertales, con una cara más grande, disfrutaban de áreas frontales más amplias que las de los HAM, aunque las diferencias no son significativas, lo que puede ser debido al tamaño limitado de la muestra.
Por lo tanto, en términos evolutivos, la fosa anterior es libre de cambiar lateralmente, ya que en el género humano las órbitas se desplazan hacia delante, y el músculo temporal es extremadamente pequeño debido a una marcada reducción de tamaño de la boca u hocico. Por el contrario, el aumento vertical de las áreas frontales se ve comprimido en estos dos taxones por una limitación biomecánica específica, y es que en los humanos modernos y los neandertales los lóbulos frontales se encuentran directamente en la parte superior del techo de la órbita. La interacción entre estos componentes anatómicos ha comenzado en los dos linajes de forma independiente hace 150-100 ka.
La encefalización de los homínidos se ha traducido en una serie de modificaciones en la bóveda craneal, base craneal, y cara. Una posición más anterior y el desarrollo lateral de la corteza prefrontal sobre los ojos, la expansión de los lóbulos temporales y la reducción del prognatismo facial, pueden perjudicar la agudeza visual, ya que limitan el desarrollo orbital y ocular. Las restricciones sobre las órbitas y los tejidos blandos de la vista, asociada con la expansión y reubicación anterior de los lóbulos frontal y temporal, podría haber sido más severa en los humanos modernos y los neandertales que en los homínidos pequeño cerebro, como resultado de la relación espacial única entre estos rasgos anatómicos en estos grupos.
Estos cambios evolutivos, además de una disminución comprobada en el volumen orbital desde el Neolítico en China, son las tendencias más importantes a tener en cuenta en la investigación de la mayor frecuencia y gravedad de ciertas formas de reducción de la agudeza visual como la miopía y el astigmatismo.
Curiosamente, disminución de la agudeza visual también es común entre las numerosas razas de perros como el Caniche Toy, Schnauzer Miniatura, Pug, Rottweiler, Collie y Labrador Retriever. Las variedades con una mayor frecuencia y mayor severidad de los defectos de refracción también tienden a ser las que han sufrido mayores fuerzas selectivas humanas en la arquitectura craneofacial, por razones funcionales y/o estéticas: se caracterizan generalmente por pedomorfismo craneofacial y poseen un neurocraneo más frontal y globular, la cara más corta y los ojos prominentes.
Mientras que el crecimiento del globo ocular va paralelo con el crecimiento del cerebro y ambos se cree que son el producto de un mismo gen pleiotrópico, la órbita crece con el resto del cráneo. Por tanto, existe una relación alométrica negativa entre el ojo y la órbita. La reducción bien documentada en tamaño humano en general y robustez que comenzó hace 12 ka actuaría para aumentar el porcentaje de la órbita ocupada por el ojo, hasta el punto de tener que competir por el espacio. La disminución observada en el volumen orbital en Asia oriental podría exacerbar una tendencia existente hacia un mayor tamaño relativo del ojo dentro de la órbita, y potencialmente ayudar a explicar la inusual alta frecuencia de la miopía en esta región geográfica.
La enfermedad de Alzheimer se caracteriza por la acumulación de placas neuríticas compuestas por depósitos extracelulares de péptido amiloide, lo que conduce a la pérdida neuronal, atrofia cortical, y el deterioro cognitivo. A pesar de que las placas son consideradas como un factor causal fundamental de la enfermedad, la cantidad de placa no se correlaciona con alteraciones cognitivas. Los ovillos neurofibrilares, con mayor frecuencia en las áreas temporales mediales, son otra característica principal neuropatológica. Debido a que la atrofia en las primeras etapas de la enfermedad, es mayor en las áreas temporales mediales, los estudios se han centrado en los lóbulos temporales. Más recientemente, sin embargo, se ha demostrado que en las primeras etapas de la enfermedad, se pueden encontrar en la corteza parietal medial alteraciones metabólicas, neuroanatómicas, y cambios histológicos.
La enfermedad de Alzheimer se sufre sólo nuestra especie. Aunque la vida útil extendida en los seres humanos puede implicar problemas de mantenimiento de la integridad del cerebro, la especificidad y la alta prevalencia en nuestra especie pueden sugerir un vínculo evolutivo entre su patología y nuestra biología cerebral. En cualquier caso la neurodegeneración en la edad avanzada no afecta la aptitud genética, ya que influye en un período que generalmente va más allá de las etapas de la reproducción humana. En términos de evolución, esta enfermedad puede ser interpretada como un caso de pleiotropía antagónica, en la que las ventajas cognitivas son pagadas con los costos de un sistema anatómico de gran alcance, caro y delicado.
La globularidad de la caja craneana, con abultamiento de la bóveda posterior, es una de las características craneales más ostensibles de los humanos modernos en comparación con los homínidos extintos y está asociada con una dilatación geométrica del hueso parietal. No tenemos ninguna evidencia de cualquier taxón extinto con un fenotipo gradual o intermedio. En África alrededor de 200-150 ka, tenemos muestras asociadas con el linaje humano moderno que muestran abombamiento parietal, mientras que otras carecen de tales características (los neandertales muestran por ejemplo lóbulos occipitales más grandes). Por lo tanto, debemos suponer que la morfología globular moderna comenzó a evolucionar en ese momento. Tales cambios espaciales en la geometría parietal están asociados con una etapa temprana postnatal en nuestra especie que está ausente en los chimpancés y los neandertales.
Desde los primeros estudios en paleoneurología, sabemos que los lóbulos parietales muestran una sorprendente variabilidad entre los homínidos intra e interespecífica. Un análisis geométrico reciente ha demostrado que las dimensiones relativas de los precúneos son en gran parte responsables de estas variaciones.
La dilatación de la región precuneal afecta a las relaciones espaciales entre el cráneo y el cerebro. Al aumentar el área parietal, el área somatosensorial se acerca al hueso frontal, y el surco perpendicular (parieto-occipital) se desplaza desde el hueso occipital. Por contraste, en los cerebros con regiones parietales más planas, el bregma se desplaza lejos de la zona central, y el surco perpendicular se acerca al lambda. Por lo tanto, en lugar de una respuesta homogénea entre los huesos y los lóbulos, tenemos un cambio en la posición relativa de estos elementos anatómicos. Con respecto a este patrón observado, el surco precentral nunca alcanza el hueso frontal, pero el lóbulo parietal puede alcanzar el hueso occipital (el surco perpendicular puede interferir en la posición de lambda).
Debido a su posición espacial, las zonas parietales profundas representan un puente estructural y funcional entre todas las otras regiones del cerebro, y por lo tanto son más sensibles a cualquier cambio o restricción. En términos de estructura, estas áreas están restringidas entre los extremos frontal y occipital, y su morfología debe ajustar de acuerdo a la disposición espacial general de la cavidad endocraneal, por lo que las variaciones se canalizan además por tensores conectivos, como el falx cerebri.
Los vasos meníngeos medios muestran una morfología compleja y reticulada sólo en los HAM, sobre todo en su superficie parietal. Teniendo en cuenta que los sistemas vasculares comparten factores morfogenéticos comunes, la complejidad de la arteria meníngea media en nuestra especie puede sugerir razonablemente que, en general, los seres humanos modernos muestran un sistema vascular endocraneal más complejo. Las redes vasculares neurocraneales (cerebral, meníngea, diploica y pericraneal) se cree que son particularmente relevantes en la termorregulación del cerebro.
El precúneo es un componente clave cuando el cerebro está en “reposo” mientras que suspende su actividad cuando el cerebro se dedica a tareas específicas. Curiosamente, también muestra niveles excepcionales de gestión de la energía. Un precúneo más próximo a la base del cerebro, como sucede en nuestra especie, puede ser susceptible a altas cargas térmicas.
Los cambios en las áreas parietales a lo largo de la evolución humana, pueden haber creado una situación de vulnerabilidad a la neurodegeneración.
Desde los primeros estudios en paleoneurología, sabemos que los lóbulos parietales muestran una sorprendente variabilidad entre los homínidos intra e interespecífica. Un análisis geométrico reciente ha demostrado que las dimensiones relativas de los precúneos son en gran parte responsables de estas variaciones.
La dilatación de la región precuneal afecta a las relaciones espaciales entre el cráneo y el cerebro. Al aumentar el área parietal, el área somatosensorial se acerca al hueso frontal, y el surco perpendicular (parieto-occipital) se desplaza desde el hueso occipital. Por contraste, en los cerebros con regiones parietales más planas, el bregma se desplaza lejos de la zona central, y el surco perpendicular se acerca al lambda. Por lo tanto, en lugar de una respuesta homogénea entre los huesos y los lóbulos, tenemos un cambio en la posición relativa de estos elementos anatómicos. Con respecto a este patrón observado, el surco precentral nunca alcanza el hueso frontal, pero el lóbulo parietal puede alcanzar el hueso occipital (el surco perpendicular puede interferir en la posición de lambda).
Debido a su posición espacial, las zonas parietales profundas representan un puente estructural y funcional entre todas las otras regiones del cerebro, y por lo tanto son más sensibles a cualquier cambio o restricción. En términos de estructura, estas áreas están restringidas entre los extremos frontal y occipital, y su morfología debe ajustar de acuerdo a la disposición espacial general de la cavidad endocraneal, por lo que las variaciones se canalizan además por tensores conectivos, como el falx cerebri.
Los vasos meníngeos medios muestran una morfología compleja y reticulada sólo en los HAM, sobre todo en su superficie parietal. Teniendo en cuenta que los sistemas vasculares comparten factores morfogenéticos comunes, la complejidad de la arteria meníngea media en nuestra especie puede sugerir razonablemente que, en general, los seres humanos modernos muestran un sistema vascular endocraneal más complejo. Las redes vasculares neurocraneales (cerebral, meníngea, diploica y pericraneal) se cree que son particularmente relevantes en la termorregulación del cerebro.
El precúneo es un componente clave cuando el cerebro está en “reposo” mientras que suspende su actividad cuando el cerebro se dedica a tareas específicas. Curiosamente, también muestra niveles excepcionales de gestión de la energía. Un precúneo más próximo a la base del cerebro, como sucede en nuestra especie, puede ser susceptible a altas cargas térmicas.
Los cambios en las áreas parietales a lo largo de la evolución humana, pueden haber creado una situación de vulnerabilidad a la neurodegeneración.












.PNG)