Esquema
- Puntas con vástago.
- América del Norte
- Complejo Nenana.
- Tradición Paleoártica Americana.
- Tradición Paleoindia del Norte.
- Tradición Paleoindia.
- Complejo Clovis o Complejo del Llano.
- Culturas del Este.
- Complejo Goshem.
- Complejo Folsom.
- Tradición Plano.
- Complejo Plainview.
- Culturas de las Grandes Llanuras del Norte.
- América Central y del Sur
- Protolítico, Lítico Inferior o Estadio Pre-puntas de Proyectil.
- Tradición Itaparica.
- Transición.
- Industria Paleoindia del Sur, Milítica o Cenolítica.
- Grupo y Horizonte Andino de Puntas Lanceoladas o Foliáceas.
- Puntas Saladillo.
- Complejo Tigre.
- Complejo Pay Paso.
 |
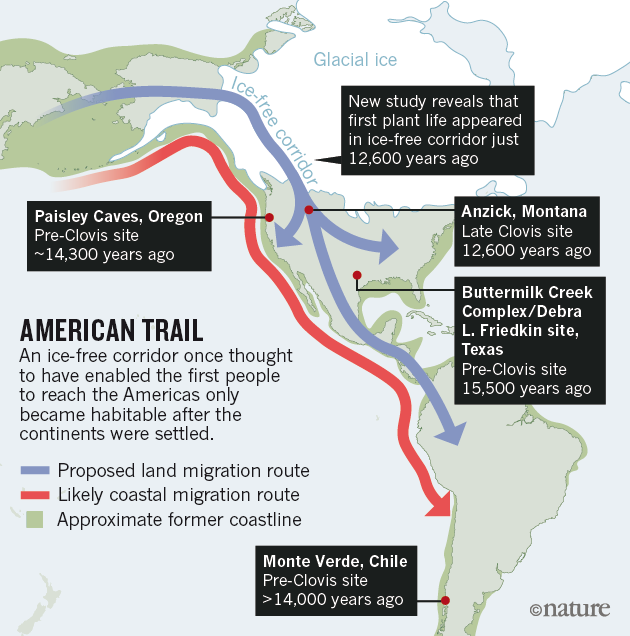
| Hallazgos líticos en América de datación más antigua. Science. |
Puntas con vástago.
Williams et al (2018) han estimado la edad de los hallazgos más antiguos del yacimiento Gault en Texas Central, en hace 20-16 ka. El conjunto refleja una tecnología de puntas de proyectil con vástago más estrecho en la parte proximal (stemmed) o vástago lanceolado así como una tradición de bifaz y de hoja y núcleo.
Davis et al (2019) han estudiado los utensilios y láminas líticos recuperados en Cooper’s Ferry, cuenca del río Columbia, Idaho, datados en hace 16,56-15,28 ka y que pudieron haber sido utilizados en el procesamiento de alimentos, asociadas a restos fósiles de caballos extintos y otros pequeños mamíferos. Tecnología de puntas de proyectil stemmed y unfluted (no acanaladas), similares a las más antiguas de la isla japonesa de Hokkaido.
Waters et al (2018) han informado del hallazgo en Debra L. Friedkin, Texas, de puntas con vástago con dataciones entre hace ca 15.500-13.500 ka, con la aparición de puntas tringulares lanceoladas hace ca 14.000 años. Las puntas lanceoladas pudieron originarse in situ a partir de las puntas con vástago o bien corresponder a una migración diferenciada.
En el congreso de la Society for American Archaeologists de 2017, Daryl Fedje presentó el hallazgo en Haida Gwaii en la Columbia Británica, Canadá, de puntas con vástago y núcleos discoides con una datación de al menos 12.700 años. Estos núcleos también están presentes en la cercana isla Triquet ocupada hace 14.000 años. Matthew Des Lauriers y Loren Davis hallaron una colección similar de núcleos en la Isla de Cedros, Baja California, donde las fechas de radiocarbono sitúan la ocupación humana en hace casi 13.000 años. En Sudámerica, Quebrada Jaguay ha deparado una colección similar y parecida datación (Sandweiss, 1998).
Davis et al (2019) han estudiado los utensilios y láminas líticos recuperados en Cooper’s Ferry, cuenca del río Columbia, Idaho, datados en hace 16,56-15,28 ka y que pudieron haber sido utilizados en el procesamiento de alimentos, asociadas a restos fósiles de caballos extintos y otros pequeños mamíferos. Tecnología de puntas de proyectil stemmed y unfluted (no acanaladas), similares a las más antiguas de la isla japonesa de Hokkaido.
Waters et al (2018) han informado del hallazgo en Debra L. Friedkin, Texas, de puntas con vástago con dataciones entre hace ca 15.500-13.500 ka, con la aparición de puntas tringulares lanceoladas hace ca 14.000 años. Las puntas lanceoladas pudieron originarse in situ a partir de las puntas con vástago o bien corresponder a una migración diferenciada.
En el congreso de la Society for American Archaeologists de 2017, Daryl Fedje presentó el hallazgo en Haida Gwaii en la Columbia Británica, Canadá, de puntas con vástago y núcleos discoides con una datación de al menos 12.700 años. Estos núcleos también están presentes en la cercana isla Triquet ocupada hace 14.000 años. Matthew Des Lauriers y Loren Davis hallaron una colección similar de núcleos en la Isla de Cedros, Baja California, donde las fechas de radiocarbono sitúan la ocupación humana en hace casi 13.000 años. En Sudámerica, Quebrada Jaguay ha deparado una colección similar y parecida datación (Sandweiss, 1998).
Fedje considera que estos hallazgos son similares a los de Siberia y Todd Braje ve similitudes a lo largo de la cuenca del Pacífico, de Japón a Perú.
Complejo Nenana
Entre 11,6-10 ka.
- Ben Potter ha comunicado en una conferencia (2015) el hallazgo en Mead de colgantes de hueso y huesos trabajados con una datación de hace unos 12,3 ka.
Yacimientos en el valle del Tanana (Walker Road, Dry Creek
component 1 y Moose Creek) y de su afluente el Nenana (Swan Point), en el
interior de Alaska. Se trata de yacimientos pequeños, localizados en riscos con
buena visibilidad, ocupados por cortos periodos de tiempo
Puntas triangulares y lanceoladas con la base cóncava,
perforadores, raspadores, raederas, buriles, percutores, yunques, cuchillos
unifaciales, lascas sin retocar y pieces
esquillées.
Probablemente se trataba de cazadores de macrofauna y caza
oportunista de pequeños animales. Caza o carroñeo de mamut.
Debido a sus parecidos, parece estar relacionado con el
Complejo Clovis.
Tradición Paleoártica Americana
10,5-8 ka.
Alaska y costa Noroeste del Pacífico.
Enraizado en las industrias de microláminas del Paleolítico
Superior Final de Siberia. Elevado número de yacimientos. Es posible que los
más antiguos estén bajo el mar. Economía costera con pesca y caza de mamíferos
marinos. Probablemente utilizaban la navegación.
Es posible que estos grupos procedan de una segunda oleada
de entrada en América.
Dixon distingue tres variantes regionales.
1. Tradición
Paleoártica Americana propiamente dicha. Costas del mar de Bering, mar de
Chukchi y Océano Ártico y entorno terrestres adyacentes. Caza de mamíferos
marinos y terrestres y pesca. Posiblemente estos grupos fueron los primeros en
introducir el arco y la flecha en Norteamérica hace 10,5 ka.
2. Complejo
Denali. Interior de Alaska. Grupos cazadores de caribú y bisonte.
3. Tradición
microlaminar de la costa noroeste. 9-8,5 ka. Archipiélago Kodiak, costa del
Pacífico del suroeste de Alaska, Columbia Británica, Washington y Oregón.
Tradición Paleoindia del Norte
10,5-8,5 ka.
Ocupa el mismo espacio cronológico y geográfico que la
Tradición Paleoártica Americana propiamente dicha y el Complejo Denali. Podría
deberse al uso de asentamientos específicos para distintas tareas. Para Dixon
se trata de dos tradiciones culturales y tecnológicas diferentes.
Puntas acanaladas. Proyectiles
TRADICIÓN PALEOINDIA
 |
| Cache sites de la cultura Clovis. J. DavidKilby y Bruce B. Huckell (2014) |
 |
| Puntas Clovis. Amick, 2016. |
 |
| Hallazgos de puntas Clovis. Amick, 2016. |
Complejo Clovis o Complejo del Llano.
Recibe su nombre de la ciudad de Clovis, al este de Nuevo México, situada en el límite occidental del Llano Estacado.
Entre ca 13,4-12,8 ka calBP en el Oeste y ca 12,8-12,5 ka calBP en el Este, aparece la punta Clovis, bastante variable.
Salvo Mason (1962), en función de las dataciones, se considera que las puntas Clovis se originaron en el Oeste, quizá derivadas del Complejo Nenana. Sin embargo, se han hallado más puntas lanceoladas y más diversas en el Este.
Entre ca 13,4-12,8 ka calBP en el Oeste y ca 12,8-12,5 ka calBP en el Este, aparece la punta Clovis, bastante variable.
La punta Clovis tiene forma lanceolada, 7-15 cm, bifacial, plana y con la base cóncava. Los costados se expanden desde la base y presentan una acanaladura basal lograda por medio de la remoción de una o varias lascas. La base acanalada de las puntas absorbe energía en el impacto y por tanto incrementa la robustez de las puntas y la capacidad de soportar el estrés físico a través de la redistribución y reubicación de los daños (Thomas et al, 2017).
La base propicia su inserción en un elemento de madera o hueso para fabricar una lanza y para ello los laterales proximales tienen unos filos menos cortantes, lo que facilita su sujeción mediante tiras de cuero. Su abundancia nos da idea de la importancia de la caza. Se ha especulado que se utilizaba de forma especializada para la caza de animales grandes, pero hay indicios de uso para la caza de lepóridos e incluso en vegetales, lo que supone una gran versatilidad.
La base propicia su inserción en un elemento de madera o hueso para fabricar una lanza y para ello los laterales proximales tienen unos filos menos cortantes, lo que facilita su sujeción mediante tiras de cuero. Su abundancia nos da idea de la importancia de la caza. Se ha especulado que se utilizaba de forma especializada para la caza de animales grandes, pero hay indicios de uso para la caza de lepóridos e incluso en vegetales, lo que supone una gran versatilidad.
Aparecen también puntas de hueso, láminas, raspadores y raederas. Son representativos los raspadores terminales con espolón, muchas veces hechos con navajas prismáticas obtenidas de núcleos poliédricos.
La técnica lítica se basa en la percusión directa de alta precisión.
Según la hipótesis de adaptación regional al entorno (Buchanan et al, 2014), los grupos Clovis adaptaron su equipamiento de caza a las características de las presas y al hábitat local. Esta adaptación regional es negada por la hipótesis de adaptación en todo el continente.La técnica lítica se basa en la percusión directa de alta precisión.
Salvo Mason (1962), en función de las dataciones, se considera que las puntas Clovis se originaron en el Oeste, quizá derivadas del Complejo Nenana. Sin embargo, se han hallado más puntas lanceoladas y más diversas en el Este.
Los yacimientos más antiguos son Aubrey (Texas) y El Fin del Mundo (Sonora; Guadalupe Sánchez et al, 2014, Guadalupe Sánchez et al, 2015) y
el más reciente Debert (Nueva Escocia). El Fin del Mundo es también el yacimiento más austral, si bien en 1993 el Proyecto Arte Rupestre Baja California Sur identificó una punta clovis en una colección particular y 1952 Aschmann comunicó el hallazgo de una punta Clovis en Rancho San Joaquín, algo más al sur (Hyland y Gutiérrez). La influencia de la cultura Clovis parece haber llegado hasta Venezuela y Ecuador.
Los yacimientos más conocidos están relacionados con la
matanza de grandes mamíferos, mamut principalmente, y se encuentran en Nuevo
México, Colorado, Arizona, Wyoming y Oklahoma. También hay hallazgos en el este
de Estados Unidos.
Para Fiedel, las bandas establecían territorios circulares
amplios con un radio aproximado de 160 km.En función de los artefactos hallados
y del entorno, se distinguen varios tipos de asentamiento:
En el oeste, Dixon distingue tres tradiciones:- Los relacionados con el abastecimiento de materias primas. Los yacimientos de Fine Mine Hall (este de Nueva York), Williamson (Virginia) y Flint Run (Virginia) muestran el aprovisionamiento de sílex, jaspe o calcedonia de gran calidad.
- Cache sites, relacionados con actividades de caza. En zonas altas, controlando ríos, lagos o valles. Ocupados por pequeños grupos a la espera del paso de los animales. Richie Roberts (Washington), Anzick (Montana), Buche Cache (Kansas), Simon Site (Idaho) y Drake Cache (Colorado). Contenían escondrijos de herramientas (bifaces, puntas de proyectil, cuchillas, núcleos, escamas, piezas de hueso y marfil, etc.). Muchas características de los Cache Clovis parecen inusuales y reflejan un alto grado de conocimiento y de apreciación estética. Incluyen materiales exóticos y en algunos caches se ha encontrado ocre rojo. En al menos cuatro caches (Anzick, East Wenatchee, Fenn y Simon) hay indicios de comportamiento ritual pero pueden representar tradiciones locales del norte de las Rocosas y del Noroeste. Con pocas excepciones, la principal diferencia entre los Cache y otros conjuntos es la inclusión de artefactos con poco uso (J. David Kilby y Bruce B. Huckell, 2014). Diversas teorías, especulan sobre su uso funcional, social y simbólico.
- Kill sites, lugares de matanza de grandes mamíferos. Cerca de lagunas o arroyos, sobre todo en el centro-oeste de Estados Unidos. Dent (Colorado), Lehner (Arizona), Blackwater Draw (Clovis, Nuevo México), Colby (Wyoming) y Hiscock (Nueva York).
- Campamentos base. De gran tamaño y gran diversidad de artefactos. No existen evidencias de estructuras, por lo que las cabañas podrían ser de ramas, musgo y piel. Debert (Nueva Escocia), Bull Brook (Massachusetts), Thunderbird (Virginia), Shoop (Pennsylvania) y West Athens Hill (Nueva York). Dixon cita una cifra entre 30-60 individuos.
- Western Fluted Points Tradition. 11,5-10,5 Tradición de puntas lanceoladas del oeste, con base cóncava. Mayor adaptación a la recolección.
- Western Stemmed Point Tradition. 11-7 ka. Tradición con puntas que tienen los laterales proximales más estrechos. Yacimientos importantes: Buhl (Idaho) y Cooper (noroeste de Oklahoma).
- San Dieguito/Sulphur Spring Complexes. 10-8 ka. Puntas alargadas y relativamente estrechas.
La transición al complejo Folsom tuvo lugar quizá en menos
de 100 años Puede que el cambio medioambiental junto con la extinción de los
mamuts obligara a un cambio en el modo de vida. Se ha especulado con la relación entre la caída de un meteorito y el evento climático Younger Drias y la desaparición de la megafauna de América del Norte y de la tecnología Clovis (Petaev et al, 2013; Moore et al, 2017).
 |
| Conjunto Clovis de East Wenatchee Cache Site |
Culturas del Este
David G. Anderson, Ashley M. Smallwood y D. Shane Miller (2015) resumen el estado actual de la arqueología paleoindia en el sureste de USA.
- Pre-Clovis o Paleoindio Temprano (hace > 13.250 años cal)
- Punta Miller.
- Punta lanceolada no acanalada, hallada por primera vez en el abrigo Meadowcroft, Pennsylvania en el Estrato IIa cuyas fronteras fueron datadas por radiocarbono en hace 13.375 ± 988 años cal y 15.389 ± 1,219 años cal. Las dataciones son dudosas.
- Punta Cactus Hill
- Punta triangular. Debe su nombre al yacimiento de Cactus Hill, Virginia, donde se hallaron dos ejemplares. Dos muestras de carbón asociadas interpretadas como restos de hogares fueron datadas en hace 18.308 ± 112 años cal y 20.428 ± 95 años cal (Feathers et al, 2006; McAvoy y McAvoy, 1997). Las dataciones son dudosas.
- Clovis o Paleoindio Medio (hace ca 13.250-12.850 años cal; Waters y Stafford, 2007).
- Disponemos de amplias evidencias de asentamientos humanos, aunque no se conoce la forma en que se extendió la cultura Clovis y las fechas fiables son escasas. La mayoría de los hallazgos vienen de contextos de superficie. Sólo en raras ocasiones se han encontrado grandes conjuntos en el contexto de excavación, por lo general relacionados con canteras, en los que la mayor parte del material recuperado está constituido por desechos de talla. A diferencia del oeste de USA, hay poca evidencia de matanza de megafauna o cachés líticos. Salvo en el norte de la región, los raspadores con mango son poco frecuentes. También es rara la evidencia de estructuras. Los restos de actividad disminuyen a medida que aumenta la distancia a las fuentes de materia prima de forma que las macrobandas dominaban un espacio hasta 300 km de las mismas.
- Johnson en Nashville, Tennessee es el yacimiento más antiguo, aunque las dataciones son dudosas (Broster et al, 2013).
- Cactus Hill en Virginia ofrece la única datación ampliamente aceptada: hace 12.786 ± 55 años cal (McAvoy y McAvoy 1997; Wagner y McAvoy 2004).
- Una varilla de marfil de Sloth Hole, Florida produjo una datación de hace 12.913 ± 74 años cal (Hemmings, 2004, 2005).
- Wittke et al (2013) y Goodyear (2013) han informado de una datación asociada con los depósitos Clovis de Topper en Carolina del Sur de hace 12.837 ± 79 años cal, pero el contexto de la fecha no se ha publicado con amplitud.
- Post-Clovis o Paleoindio Tardío (hace ca 12.850-11.700 años cal, coincidiendo con el Dryas Reciente; Bousman y Vierra, 2012; Fiedel, 2015; Walker et al, 2009).
- Se produce un gran cambio climático y cultural, con grandes extinciones y reubicaciones de flora y fauna, transgresiones marinas y reversiones, y cambios en los regímenes fluviales. La regionalización cultural había comenzado en el Paleoindio Medio (Smallwood 2012; Smith et al 2015; Thulman 2006), y es muy evidente en el Paleoindio Tardío, representada por un aumento en el número y diversidad de los tipos de puntas estriadas y no acanaladas. Se desconoce la causa de esta variación.
- Disminuye el número de yacimientos y hallazgos, lo que se interpreta como una reorganización o disminución de la población, especialmente en el norte de Alabama y el sur de Virginia. Las extinciones de la megafauna habrían dado lugar a cambios en la dieta y a su vez en la tecnología, con aumento de formas entalladas y reafiladas, tal vez relacionadas con la caza menor.
- El inicio del periodo está representado por formas de punta Cumberland y Redstone, derivadas de la punta Clovis (Goodyear 2006, 2010) y ocasionalmente, tipos de las llanuras, similares a Folsom. Goodyear (2006) propone que el uso de la percusión indirecta permitió un mayor control del acanalamiento y que las puntas fueron diseñadas para penetrar, mientras que las Clovis estaban pensadas para cortar, un diferente propósito funcional que se asocia con cambios en las comunidades de fauna. Los yacimientos correspondientes no se han fechado con seguridad.
- Cumberland
- El canal se extiende desde la base hasta la punta, con una cintura de enmangamiento y una base ligeramente cóncava. Los bordes se caracterizan por su talla fina por presión marginal (Lewis, 1954). Gramly (2008, 2009, 2012), sostiene que el tipo Cumberland es pre-Clovis. Aparece en el Medio Sur.
- Redstone
- Puntas triangulares grandes y medianas con concavidades basales profundas y largos canales múltiples en ambas caras, aunque a veces son cortos; la hoja es recta y termina con una punta distal aguda (Daniel y Goodyear, 2006; Goodyear 2006; Mahan 1964). Aparece en la parte oriental del sudeste de USA.
- Las puntas Folsom aparecen al este, hasta Illinois e Indiana (Munson 1990). Similares a Folsom:
- Sedgwick, en Arkansas (Morse y Morse, 1983).
- Barnes en el Medio Oeste.
- Northumberland en Pennsylvania.
- Michaud-Neponset en Nueva Inglaterra y marítimas.
- En algún momento durante la primera parte del Paleoindio Tardío, desaparecen los acanalamientos en todo el sudeste y la variación morfológica aumenta, incluyendo los tipos:
- Beaver Lake y Quad en todo el sudeste.
- Arkabutla, Coldwater, y Hinds en Mississippi.
- San Patrice principalmente en el trans-Mississippi al sur de Luisiana y zonas adyacentes.
- Suwannee en Florida y la llanura costera de Georgia y Carolina del Sur.
- Este reemplazo sólo se ha insinuado estratigráficamente en unos pocos yacimientos, en particular Dust Cave in Alabama (Driskell 1994; Sherwood et al, 2004), Hester en Mississippi (Brookes, 1979), y Silver Springs en Florida (Dunbar 2006c; Neill 1958). En Dust Cave se hallaron componentes Quad y Beaver Lake por debajo de componentes Dalton (Driskell 1994; Sherwood et al, 2004).
- La datación de las puntas Dalton es incierta, pero parece abarcar la parte más tardía del Dryas Reciente y posiblemente el Holoceno temprano (Driskell et al 2012; Ellis et al, 1998; Goodyear 1982; Lopinot et al 1998, 2000; Miller y Gingerich, 2013a , 2013b; Morse 1997b ;Morse et al, 1996). Bradley (1997) ha argumentado la posibilidad de que las puntas Dalton apareciesen antes, evolucionando directamente de Clovis en el valle del Mississippi central, donde se han observado acanalamientos en algunas puntas de Sloan en Arkansas noreste. Las puntas Dalton se han recuperado en grandes cantidades en todo el sudeste de USA, salvo en Florida, donde son sustituidas por las puntas Suwannee y formas similares (Dunbar 2006c. Dunbar y Hemmings, 2004. Se han propuesto modelos de asentamiento y subsistencia de las poblaciones Dalton con movimiento desde bases centrales hacia sitios con actividades especiales (incluidas las canteras), y cementerios alejados de los sitios residenciales (Morse 1971, 1973, 1975a; Morse y Morse 1983; Gillam, 1996a, 1999). Se observa la explotación de una amplia gama de flora y fauna, con énfasis en la caza de ciervos de cola blanca (Goodyear 1974; Morse 1971, 1973, 1997a , 1997b). Debido a la gran cantidad de sitios y a las características inusuales observadas en algunos de ellos, puede hablarse de un florecimiento cultural en el Valle Central del Mississippi. Sitios como Lace y Brand, parecen haber sido lugares de asentamiento prolongado, mientras que en Sloan han aparecido cementerios señalizados, denotando posible territorialidad o el control de ciertas áreas o recursos (Kelly 2013). Las puntas Sloan son grandes bifaces que reflejan algún tipo de ceremonialismo y aparecen solas y en cachés en el Valle Central del Mississippi. Walthall y Koldehoff (1998) proponen la existencia de una red ceremonial o de interacción entre los pueblos de la zona y Anderson (2002) y Sassaman (2010) una compleja sociedad de cazadores-recolectores.
- Para la fase media del Dryas Reciente y la transición al Holoceno temprano, hay fechas disponibles probablemente debido al aumento de uso de cuevas y abrigos rocosos, que proporcionan un mejor contexto para la preservación.
- Dust Cave es el yacimiento clave para el establecimiento de la relación cronológica entre Quad / Beaver Lake, Dalton, y puntas tempranas con muescas laterales (Sherwood et al, 2004), demostrando que las puntas con muescas se extienden durante el límite entre el Dryas Reciente y el Holoceno.
- En Smith Mountain, Virginia (Childress y Blanton, 1997) se recuperó un bifaz tipo Plano. Una muestra de carbón produjo una fecha de hace 11.790 ± 165 años cal.
Complejo Goshem
11-10,5 ka.
George Frison e Irwin-Williams han sugerido que puede ser
una cultura de transición entre Clovis y Folsom.
·
A favor de este planemiento está el
descubrimiento en el yacimiento de Hell Gap (Wyoming) por debajo de un nivel
Folsom, al igual que en Mill Iron (Montana) y Carter/Kerr-McGee (Wyoming).
·
En contra, es difícil explicar por qué se
pasó de puntas ligeramente acanaladas
tipo Clovis a las no acanaladas para volver otra vez a las acanaladas tipo
Folsom.
Las puntas Goshem son puntas Clovis sin acanalamiento. Puede
que constituyan una simple variante no acanalada.
 | ||||
| Ejemplos de puntas lanceoladas de Norteamérica: (a) Clovis (Logan Co., Kentucky); (b) Dalton (Lyon Co., Kentucky); (c) Folsom (Roosevelt Co., New Mexico); (d) Cumberland (Colbert Co., Alabama); (e) Gainey/Bull Brook (Essex Co., Massachusetts); (f) Crowfield (Addison Co., Vermont) |
 |
| Hallazgos de puntas Folsom. Amick, 2016. |
Complejo Folsom
10,9-10,2 ka.
Desde las Montañas Rocosas a las Grandes Llanuras y desde
Dakota del Norte al norte de México. En el Lejano Oeste, esta tradición es
coetánea de la Western Stemmed Point Tradition.
El primer descubrimiento fue realizado en 1908 por George
McJunkin cerca de Folsom (Nuevo México).
Otro yacimiento importante es Lindenmeier (norte de
Colorado), datado en 10.660 años. Excavado en los años 30 y estudiado
nuevamente en los 70. Campamento base. Aparecieron instrumentos líticos de
obsidiana de una fuente a 560 km al noroeste. Se localizaron numerosas agujas
de hueso.
Más yacimientos: Indian Creek (Montana), Cooper (Oklahoma) y
12 Mile Creek (Kansas).
Además de puntas, en los yacimientos aparecen lascas,
cuchillos bifaciales muy delgados, perforadores, raspadores, raederas,
choppers, piedras abrasivas, pigmentos y piedras para machacar pigmentos.
Por lo general, las puntas Folsom son más pequeñas que las
Clovis, tienen la base cóncava y cuentan con un acanalado más extenso, que se
aproxima al extremo final del artefacto. Son objetos de excepcional calidad
técnica, fabricados con excelente materia prima, de grano muy fino.
El canal que presentan las puntas Folsom se obtuvo, según
Donald Crabtree, desgajando las lascas presionándolas hacia abajo con una
muleta en forma de T apoyada contra el pecho. La punta de la muleta se presiona
en el punto exacto del núcleo mientras se sujeta éste con firmeza. La mejora
funcional con respecto a las Clovis, es objeto de discusión.
Extinguido el mamut, los Folsom cazaban bisontes, de mayor
tamaño que los actuales. Hay evidencias de matanzas sistemáticas en Stewart’s
Cattle Guard (sur de Colorado) y Agate Basin (Wyoming). Hay indicios de que los
excedentes se congelaban. Además del bisonte, los Folson explotaban los
recursos de forma generalista. Se han encontado hogares con restos de ceniza.
Es posible que utilizaran tiendas similares a las de los nativos recientes. Los
campamentos estaban situados en lugares altos, buenos puntos de observación.
 |
| Puntas Clovis y Folsom |
Tradición Plano
10,2-8 ka.
La tradición incluye varios tipos de punta, con diferencias
regionales y temporales.
El cambio de Folsom a Plano parece que fue bastante abrupto.
El animal más cazado continúa siendo el bisonte, y las
estrategias de caza son similares a las del Complejo Folsom, aprovechando la
topología del terreno para crear trampas naturales, sin descartar la
utilización de vallados de troncos. Es posible que se congelasen o ahumasen los
excedentes. También hay evidencias de piedras de moler, por lo que además de la
caza podemos suponer estrategias de recolección.
Quiza fueron cambios en los patrones de asentamiento a lo
largo del flanco este de las Montañas Rocosas los que dieron lugar a la
Tradición Arcaica que aparece en las regiones áridas del oeste y en los bosques
orientales de Norteamérica, ligada a estrategias de caza oportunista de
pequeños mamíferos y recolección y menos dependiente del bisonte.
La Tradición Plano desapareció coincidiendo con un cambio
climático que dio lugar a un aumento de la sequía (periodo Altitermal). Terminó
así la época de las grandes cacerías comunales. La disminución de recursos
conllevó la de la de la población. Pero la escasez de yacimientos puede deberse
también a la gran erosión que caracteriza ese periodo.
Yacimientos:
Yacimientos:
- North Fork, Kelly Forks, Clearwater River. Se hallaron 19 puntas, la más antigua datada en 11 ka. El asentamiento fue usado de forma regular por los antiguos cazadores-recolectores, principalmente para la fabricación de herramientas y procesamiento. Se ha comprobado el uso de herramientas para el procesamiento de conejos.
Complejo Plainview
10-8 ka.
En las grandes llanuras del sur.
Son equiparables a las puntas Goshem.
Recibe su nombre del yacimiento de Texas excavado en 1944.
Las puntas Plainview son lanceoladas con base cóncava,
similares a las Folsom pero sin acanaladura
Para Hofman y Graham (1998) pudo ser una fase intermedia
entre el Complejo Folsom y el Complejo Cody.
En el yacimiento Olsen-Chubbock (Colorado), datado en 10,15
ka, excavado entre 1958-60 por Joe Ben Wheat aparecieron restos de 190 bisontes
asociados a 27 puntas además de otros artefactos. Muchos animales no fueron
consumidos. Se calcula que en la matanza y consumo estuvieron implicadas
150-200 personas. Las puntas son de dos tipos, por lo que es posible que
participasen don bandas.
·
Puntas Firstview, lanceoladas, con base recta.
·
Puntas San Jon.
Culturas de las Grandes Llanuras del Norte
Incluye los complejos:
·
Agate Basin. 10,5-10 ka. Puntas lanceoladas,
alargadas, con un ligero estrechamiento en la zona proximal. Más estrechas y
largas que las puntas Clovis y Folsom. En Agate Basin estas puntas aparecieron
por encima de un nivel Folsom. Se hallaron puntas similares en Packard (noreste
de Oklahoma) y en Canadá. Para Stanford, pudo tener su origen en el Lejano
Oeste ya que es allí donde aparecen antes puntas similares. También es posible
que proceda del complejo Denali. En el enterramiento de Gordon Creek (norte de
Colorado) aparecieron restos humanos y artefactos cubiertos de ocre rojo, lo que
podría implicar algún tipo de ritual relacionado con la muerte.
·
Hell Gap. 10,2-9,5 ka. Recibe su nombre del
yacimiento del sureste de Wyoming. Parece un desarrollo del Agate Basin. Las
puntas son más estrechas en la parte mesial proximal y tienen una morfología
algo más ovoide que las Agate. El yacimiento de Jones-Miller (noreste de
Colorado) ha sido interpretado como un lugar de matanza de bisontes. Para
Stanford, algunos hallazgos sugieren rituales asociados a matanzas comunales.
En Casper (Wyoming) fueron cazados 100 bisontes.
· Alberta. 9,8-9,2 ka. Reconocido por primera vez
en el sur de Alberta. Puntas con pedúnculo. El yacimiento de Horner (noroeste
de Wyoming) ofrece puntas de tipo Cody y de tipo Alberta, por lo que ha sido
interpretado como una transición entre ambos complejos. En Hudson Meng
(Nebraska) se localizaron restos como mínimo de 500 bisontes asociados a 21
puntas de tipo Alberta y a algún cuchillo de tipo Cody.
· Cody.9,5-8,4 ka. Identificado a partir de los
hallazgos en Horner (noroeste de Wyoming). Para Dixon, este complejo evolucionó
a partir del Alberta. Algunos autores sugieren una conexión con el Complejo
Plainview. Puntas alargadas y muy estrechas, cuya zona proximal es ligeramente
más estrecha. Mucha variabilidad. Los tipos de puntas más significativos son:
o
Las puntas Eden (yacimiento al oeste de Wyoming)
o
Puntas Scottsbluff (yacimiento al oeste de
Nebraska)
Paleoindios de América Central y del Sur.
Cazadores de grandes mamíferos, posiblemente incrementaron
la proporción de caza menor al desaparecer la megafauna pleistocénica. Hay
evidencias de una rápida domesticación del perro. Después de hace 11 ka los
vegetales adquirieron mayor relevancia en la alimentación. En muchos casos, se
practicaba la trashumancia. En las zonas costeras se explotan los recursos
marinos de forma especializada.
La ocupación humana debió tener lugar mucho antes de la
época Clovis.
Se observa una gran variedad en la morfología de las
herramientas líticas. Schöbinger (1988) distingue dos etapas y una época
transición:
PROTOLÍTICO, LÍTICO INFERIOR O ESTADIO PRE-PUNTAS DE PROYECTIL.
Industrias de guijarros y lascas.
Noreste y este de Brasil.
Tradición Itaparica
 |
| Principales yacimientos del centro y norte de Brasil, anteriores a hace 8 ka cal. Los círculos negros indican sitios donde al menos una capa proporcionó artefactos unifaciales. (1) Santa Elina. (2) MT-SL-31 (Mato Grosso). (3) GO-JA-01. (4) GO-JA-02. (5) GO-JA-03. (6) GO-JA-14. (7) GO-JA-26 (Goiás). (8) MS-PA-02 (Mato Grosso do Sul). (9) Lapa do Gentio II. (10) Lapa da Foice I. (11) Lapa do Dragão. (12) Lapa do Boquete. (13) Lapa do Bichos. (14) Lapa do Boqueirão Soberbo. (15) Lapa Pequena. (16) Santana do Riacho- (17) Lapa Vermelha IV. (18) Cerca Grande VI. (19) Lapa das Boleiras. (20) Lapa do Santo (Minas Gerais). (21) Abrigo do Pilão (Bahia). (22) Toca do Boqueirão da Pedra Furada. (23) Toca do Pica-Pau. (24) Toca do Sítio do Meio. (25) Toca do Bojo I. (26) Toca do Baixão do Perna I. (27) Toca dos Coqueiros. (28) Toca da Baixa das Cabaceiras. (29) Toca da Cerca do Elias. (30) Toca do João Leite. (31) Toca do Pau Dóia. (32) Toca do Paraguaio. (33) Toca Nova do Inharé (Piauí). (34) Miracema 1. (35) Miracema 2. (36) Lajeado 18. (37) Mares 2. (38) Capivara 5 (Tocantins). (39) Justino (Sergipe). (40) Gruta do Padre. (41) Furna do Estrago. (42) Pedra do Caboclo. (43) Chã do Caboclo (Pernambuco). (44) Pedra do Alexandre. (45) Mirador (Rio Grande do Norte). (46) Lapa do Sumidouro. (47) Vale da Pedra Furada. (48) Toca da Tira Peia. |
TRANSICIÓN
Comprende los yacimientos entre 13-11 ka con puntas
triangulares o levemente amigdaloides. Se han encontrado puntas en yacimientos
de Centroamérica (Fell en Panamá, Turrialba en Costa Rica) que podrían
confirmar el nexo con las puntas Clovis, pero estos yacimientos son de difícil
datación.
Yacimientos de Los Toldos, El Ceibo y Río Pinturas II y III
(Patagonia). Además de industria lítica, aparece arte rupestre de puntos y
elementos geométricos simples, figuras de guanacos con cazadores esquematizados
y manos en negativo.
El fuerte parecido entre los hallazgos de Costa Rica y
Patagonia indica una rápida ocupación humana (Fiedel).
INDUSTRIA PALEOINDIA, MILÍTICA O CENOLÍTICA.
- Oeste: Yacimientos de Cueva Fell (Chile) y El Inga (Ecuador).
- Este y sur de Brasil, Uruguay, Argentina y Patagonia.
Las puntas de cola de pescado muestran dos variantes principales (Suárez 2001, 2006).
- El estilo clásico, con un tallo marcado y hombros redondeados (ángulo del hombro-tallo entre 120º-160°).
- Otra variante, con hombros muy marcados (ángulo entre 90°-110°).
Grupo y Horizonte Andino de Puntas Lanceoladas o Foliáceas.
11-6 ka.
- Complejo Paiján. 11-10 ka. Localizado en la costa norte y central de Perú con posibles ramificaciones en la sierra ecuatoriana. Caracterizado por puntas pedunculadas.
- Complejo surandino de Tuina-Inca Cueva 4. 10,8-9 ka. Puntas triangulares delgadas, de tamaño mediano a pequeño. Pinturas rupestres geométricas.
- Complejo Paiján. 11-10 ka. Localizado en la costa norte y central de Perú con posibles ramificaciones en la sierra ecuatoriana. Caracterizado por puntas pedunculadas.
- Complejo surandino de Tuina-Inca Cueva 4. 10,8-9 ka. Puntas triangulares delgadas, de tamaño mediano a pequeño. Pinturas rupestres geométricas.
Puntas Saladillo
Variante con retoque unifacial.
Complejo Tigre
Complejo Tigre
El complejo cultural Tigre aparece entre hace 12.552-11.213 años cal en los primeros componentes de Pay Paso 1, los niveles antiguos de K87 y en Laguna Canosa con una cronología radiocarbónica similar. El kit de herramientas incluye puntas de proyectil con tallo, bifaces ovalados y asimétricos de tamaño mediano, probablemente usados como herramientas de corte, grandes preformas, hojitas de corte y una variedad de raederas. Pay Paso 1 se encuentra en las riveras del Cuareim; K87 y Laguna Canosa, en las llanuras del río Uruguay (Suárez, 2015).
- La punta Tigre (Suárez 2011a) tiene un tallo ancho con lados rectos o ligeramente convexos, los hombros con muescas muy pronunciadas (ángulo ca 70-90°), una cuchilla triangular, una base convexa por retoque, adelgazamiento bifacial completo, y abrasión en los bordes del tallo. Se han hallado decenas de puntas Tigre en contextos de superficie, en Uruguay. Es importante tener en cuenta que los fabricantes de puntas de cola de pescado de Uruguay no utilizaron madera silicificada, pero sí lo hicieron los de puntas Tigre.
El complejo cultural Tigre aparece entre hace 12.552-11.213 años cal en los primeros componentes de Pay Paso 1, los niveles antiguos de K87 y en Laguna Canosa con una cronología radiocarbónica similar. El kit de herramientas incluye puntas de proyectil con tallo, bifaces ovalados y asimétricos de tamaño mediano, probablemente usados como herramientas de corte, grandes preformas, hojitas de corte y una variedad de raederas. Pay Paso 1 se encuentra en las riveras del Cuareim; K87 y Laguna Canosa, en las llanuras del río Uruguay (Suárez, 2015).
- La punta Tigre (Suárez 2011a) tiene un tallo ancho con lados rectos o ligeramente convexos, los hombros con muescas muy pronunciadas (ángulo ca 70-90°), una cuchilla triangular, una base convexa por retoque, adelgazamiento bifacial completo, y abrasión en los bordes del tallo. Se han hallado decenas de puntas Tigre en contextos de superficie, en Uruguay. Es importante tener en cuenta que los fabricantes de puntas de cola de pescado de Uruguay no utilizaron madera silicificada, pero sí lo hicieron los de puntas Tigre.
Complejo Pay Paso
El complejo cultural Pay Paso aparece en el Componente 3 de Pay Paso y en K87 por encima del complejo Tigre (entre 11.081-11.024 años cal). Se recuperaron puntas Pay Paso también en otros sitios arqueológicos a lo largo de los cursos medios de los ríos Uruguay y Cuareim. Estos tipos de puntas aparecen asimismo en las colecciones arqueológicas del curso medio del río Negro y en los valles del río Tacuarembó Grande. Además de las puntas, son características las hojitas y los raspadores (Suárez, 2015).
- La punta Pay Paso se caracteriza por un tallo corto, profundamente cóncavo en su base, bordes cóncavos divergentes en expansión hacia la base, abrasión de los bordes del tallo, hoja con bordes convexos o rectos, retoque laminar regular de la hoja, y adelgazamiento basal muy cuidadoso del vástago con cicatrices de escamas triangulares y cortas.
El complejo cultural Pay Paso aparece en el Componente 3 de Pay Paso y en K87 por encima del complejo Tigre (entre 11.081-11.024 años cal). Se recuperaron puntas Pay Paso también en otros sitios arqueológicos a lo largo de los cursos medios de los ríos Uruguay y Cuareim. Estos tipos de puntas aparecen asimismo en las colecciones arqueológicas del curso medio del río Negro y en los valles del río Tacuarembó Grande. Además de las puntas, son características las hojitas y los raspadores (Suárez, 2015).
- La punta Pay Paso se caracteriza por un tallo corto, profundamente cóncavo en su base, bordes cóncavos divergentes en expansión hacia la base, abrasión de los bordes del tallo, hoja con bordes convexos o rectos, retoque laminar regular de la hoja, y adelgazamiento basal muy cuidadoso del vástago con cicatrices de escamas triangulares y cortas.