 |
| Yacimientos más antiguos de América del Sur |
Se ha comunicado el hallazgo en la cueva Cacao 1A (Catamarca, Argentina) de dos mechones de pelo humano, herramientas de piedra tallada utilizadas para corte y raspado, unos pendientes de cobre y excrementos y fragmentos de huesos de animales que formaron parte de megafauna ya extinta. La datación mediante carbono 14 es de hace 40 ka. A la espera de dataciones complementarias.
Algunos de los yacimientos más antiguos se encuentran en América Latina, pero están sujetos a polémica (Monte Verde, Chile, antes de 25 ka, Dillehay et al, 2015; Tlapacoya, México, 24 ka; Toca da Tira Peia, Piauí, Brasil 22 ka; Arroyo del Vizcaíno, Uruguay 30-27 ka; Taima Taima, Coro Venezuela 14,44 ka; Pikimanchay, Ayacucho, Perú, 20 ka; Pedra Furada, Serra da Capivara, este de Piauí, Brasil, 32,16 ka y 20 ka; Toca do Serrote das Moendas, São Raimundo Nonato, Piauí, Brasil, 29-24 ka; Pilauco, Osomo, Chile, 15,6; Abrigo de Toca do Sitio do Meio, Serra da Capivara, este de Piauí, Brasil, 14,3 ka; Arroyo Seco 2, Tres Arroyos, Pampa, Argentina, 14 ka). En Alaska y Canadá existen lugares con restos fechados hasta en 27 ka (Bluefish II, 24,82 ka; Swan Point, Little John, Broken Mammoth y Mead; Easton et al, 2013; Goebel et al, 2008; Hoffecker y Elias, 2003), pero los datos fiables no van más allá de los 16,5-13,9 ka. Cooper's Ferry, cuenca del río Columbia, Idaho, entre hace 16,56-15,28 ka. Rimrock Draw Rockshelter, en Riley, Oregón, con más de 15,8 ka; Debra L. Friedkin, Texas, 15,5 ka. Page-Ladson, Florida, 14,55 ka; Paisley Caves, Oregón, 14,5 ka; la cueva de Wilson Butte, en Idaho, con 14,5-13 ka; Schaefer y Hebior, Wisconsin ca 14,8-14,2 ka; Triquet Island, Canadá, 14 ka; la cueva de Fort Rock, en Oregón, de 13,25 ka y otros yacimientos de Pennsylvania y Florida con unas edades de 12 ka. En 2011 se dató en 13,8 ka una punta de lanza hallada en el fósil de una costilla de mastodonte de Manis (Washington) y en 13,2 un conjunto de más de 15.000 artefactos de piedra hallados en el centro de Texas. En 2014 se dató en 22 ka un cráneo de mastodonte hallado en 1974 en la Bahía de Chesapeake junto a dos herramientas líticas, pero no está claro que estén asociados (Dennis Stanford et al, 2014). En 2015, Michael R. Waters et al dataron los restos de Wally's Beach, Canadá, de siete caballos y un camello luego de ser descuartizados, así como 29 núcleos y lascas no diagnosticables, obteniendo una fecha media de 13,3 cal ka. Los animales fueron sacrificados en un periodo corto.
Para Stuart Fiedel, no hay evidencias de intervención humana en los ecosistemas, anteriores a la cultura Clovis.
 |
| El poblamiento de América a la luz del genoma Anzick 1 |
Fósiles más antiguos (más de 10 ka).
- Hoyo Negro, un pozo profundo dentro del sistema de cuevas Sac Actun, Península de Yucatán (México). Naia; en griego, ninfa del agua. Esqueleto casi completo de una adolescente de unos 15-16 años con un cráneo intacto y DNA preservado hallado en 2007 por el equipo de Alberto Nava (National Geographic). Muestra caracteristicas craneofaciales paleoamericanas: rostro estrecho con los ojos muy separados y frente prominente y baja; nariz baja y plana; y los dientes proyectados hacia fuera. Presenta signos de caries dental y osteoporosis. La talla es pequeña (1,5 m) y la constitución frágil. Su ADN-mt pertenece al haplogrupo D1, que se cree procede de Beringia, y es exclusivo actualmente de los nativos americanos incluyendo sudamérica. 13-12 ka. James C. Chatters et al, 2014.
- Upward Sun River, Valle del Río Tanana, Alaska Central. 11,5 ka. Potter et al (2011). Potter et al (2014). Tackney et al (2015). En 2010 fueron hallados los restos de un niño enterrado de tres años de edad. En 2013, los restos de otros dos bebés enterrados, cubiertos de ocre rojo junto con cuatro piezas de cornamenta decoradas, dos puntas líticas de flecha y bifaces.
- El análisis del ADN-mt mostró haplotipos diferentes, presentes aunque raros en las poblaciones americanas actuales pero austentes en las asiáticas.
- Moreno-Mayar et al (2018) obtuvieron el genoma completo de USR1, un bebé de seis semanas. De acuerdo con los resultados, este individuo perteneció a una población diferente a la de los nativos americanos actuales y antiguos cuyo genoma se ha secuenciado hasta la fecha.
- Witt Site (Tulare Lake, CA). 11,38 ka. Willig (1991).
- Peñón III, Ciudad de México. 1959. Esqueleto parcial de una mujer de unos 25 años. 10,75. González et al (2003, 2006). Jiménez López et al (2006).
- Abrigo de Anzick, oeste de Nevada. 1968. Restos de un niño de 18 meses. 10.705 ± 35 14C años PB (aproximadamente 12.707-12.556 cal BP). Owsley y Hunt (2001). Asociado con herramientas Clovis.
- Buhl, sur de Idaho. Restos de mujer de 17-21 años y 1,65 cm de estatura. El esqueleto estaba removido y parece que fue desarticulado antes de ser enterrado. 10,675. Green et al (1998).
- Wilson Leonard, TX. 10,5 ka. Steele (1998).
- Chimalhuacán, México. 10,5 ka. González-José et al (2008).
- Mostin, norte de California. 10,47 ka. Taylor et al (1985).
- Warm Mineral Springs, Sarasota County, oeste de Florida. Restos de un varón de 30-40 años, una mujer y un individuo de 6 años. 10,26 ka. Clausen et al (1975).
- Arch Lake, NM. 10,22 Owsley et al (2010).
- Tlapacoya I, Ciudad de México. 1968. Cráneo de un varón de 30-35 años. 10,2 ka. González et al (2003, 2006).
- Marmes Floodplain, sureste de Washington. Restos quemados de 10 individuos. 10,13 ka. Sheppard et al (1987).
- Arlington Springs, Isla de Santa Rosa, sur de California. 1959. Dos fémures y un húmero. 10,08 ka. Johnson et al (2002).
América del Sur.
- Lapa Vermelha IV, Lagoa Santa, Minas Gerais, Brasil. 1975. Luzia, restos de mujer joven, de 1,5 m. Según Walter Neves (2003, 2004) presenta rasgos negroides. 11,68 ka. Prous y Fogaça (1999).
- Cerca Grande 6 y 7. Lagoa Santa, Minas Gerais, Brasil. 11 ka. Neves et al (2004, 2005).
- Pampa de Fósiles 13. Perú. 10,25 ka. Chauchat (1998).
- Sueva 1, Colombia. 10,09 ka. Correal Urrego y van der Hammen (1979).
Caribe.
- La presencia humana en Cuba data de 10-8 ka.
- Es posible que las islas del Caribe fuesen colonizadas por diversas culturas simultáneamente, procedentes de Venezuela (saladoides) y de los Andes bolivianos (huecoides), con costumbres muy diferentes.
 |
| Inferencias demográficas a partir de USR1. No se ha podido determinar el lugar de residencia de la población fundadora de todos los nativos americanos. La población que habitó en Beringia fue sustituida por otras poblaciones relacionadas. Moreno-Mayar et al (2018). |
Beringian standstill hypothesis
El término “Beringia” fue acuñado en 1937 por el botánico sueco Eric Hultén. Beringia se extiende desde las montañas Verkhoyansk en Siberia hasta el río Mackenzie en Canadá. Incluye un puente de tierra sobre el estrecho de Bering (Watson, 2017).
Emőke Szathmáry et al (1978) propusieron que las similitudes genéticas y físicas entre los dos grupos de nativos americanos existentes hoy en día surgieron cuando diferentes poblaciones fundadoras intercambiaron genes en Beringia. Según la hipótesis Standstill of Beringia (Tamm et al, 2007) Out of Beringia o Statu Quo Beringia (estancia en Beringia) una población nómada procedente del norte de Asia se asentó en Beringia hace ca 30-18,4 ka, coincidiendo con el Último Glacial Máximo (LGM), que pudo obligar a las poblaciones siberianas a buscar refugios más al sur mientras que otros grupos permanecían aislados en zonas libres de hielo en Beringia ante la imposibilidad de retirarse más al sur. Solo pudieron reanudar su camino tras el inicio del deshielo y la apertura de las rutas costeras. Graf y Buvit (2017) consideran que el origen de esta población puede no estar en Beringia, sino en el Lejano Este de Rusia o en las Paleo-Sakhalin-Hokkaido-Kuril.
Según John F. Hoffecker, Scott A. Elias y Dennis H. O'Rourke (2014), Hoffecker et al (2016) el centro de Beringia mantuvo una tundra arbustiva con algunos árboles durante el último máximo glacial y se caracterizó por temperaturas sorprendentemente suaves, dada la alta latitud. La suavidad del clima fue causada por los patrones de circulación del Pacífico Norte que llevaron aire húmedo y relativamente cálido. Una población de unas decenas de miles de personas se estacionó en el centro de Beringia durante 2,4-9 ka (Llamas et al, 2016).
Una clave para la ocupación prolongada puede haber sido la utilización de la madera como combustible. Los experimentos han demostrado que es necesaria alguna madera para utilizar el hueso como combustible. Esta población entró en las Américas a través de una ruta costera hace ca 16,0 ka, cuando quedó disponible un camino viable (Lesnek et al, 2018).
De acuerdo con evidencias genéticas, la población fundadora fue de unos pocos cientos de individuos (Hey, 2005; Fagundes et al, 2007; Kitchen et al, 2008; Gutenkunst et al, 2009; Ray et al, 2010; Wall et al, 2011; Gravel et al, 2013; Fagundes et al, 2018).
De acuerdo con las inferencias demográficas a partir del genoma completo de USR1 (Moreno-Mayar et al, 2018), los primeros pobladores de Beringia, fueron sustituidos por poblaciones relacionadas.
El tiempo en alcanzar el Sur del continente fue de ca 1,4 ka, ya que Monte Verde, en el sur de Chile, se ha datado en ca 14,6 ka.
Emőke Szathmáry et al (1978) propusieron que las similitudes genéticas y físicas entre los dos grupos de nativos americanos existentes hoy en día surgieron cuando diferentes poblaciones fundadoras intercambiaron genes en Beringia. Según la hipótesis Standstill of Beringia (Tamm et al, 2007) Out of Beringia o Statu Quo Beringia (estancia en Beringia) una población nómada procedente del norte de Asia se asentó en Beringia hace ca 30-18,4 ka, coincidiendo con el Último Glacial Máximo (LGM), que pudo obligar a las poblaciones siberianas a buscar refugios más al sur mientras que otros grupos permanecían aislados en zonas libres de hielo en Beringia ante la imposibilidad de retirarse más al sur. Solo pudieron reanudar su camino tras el inicio del deshielo y la apertura de las rutas costeras. Graf y Buvit (2017) consideran que el origen de esta población puede no estar en Beringia, sino en el Lejano Este de Rusia o en las Paleo-Sakhalin-Hokkaido-Kuril.
Las pruebas arqueológicas que apoyan este modelo, son limitadas, sobre todo en el lado canadiense.
- Yana RHS (Rhinoceros Horn Site), datado en hace 32 ka (Pitulko et al, 2004).
- Otros yacimientos de una antigüedad similar en Siberia central (Nepa, Alekseevsk) y en la parte europea de Rusia (Mamontovaya Kurya, Byzovaia).
- En unos depósitos junto al río Inmachuk (noroeste de Alaska), se encontró un fragmento de colmillo de mamut con marcas datado hace ca 40 ka, aunque lo más probable es que el colmillo fuera recogido aún fresco tras el deshielo y fuera trabajado por habitantes más recientes de la zona (Gelvin-Reymiller et al, 2006).
- Cuevas Bluefish (Yukón, Canadá), donde se han recuperado unos huesos con marcas de corte datados hace al menos 24 ka.
Los restos humanos más antiguos hallados en Siberia corresponden al yacimiento de Mal’ta (MA-1), datado en hace 24 ka. Los fósiles de Tianyuan y Yamashita-cho (Este de Asia) se han datado entre hace 42-36 ka.
La ausencia de yacimientos puede deberse entre otras causas, a la subida del nivel del mar.
Una clave para la ocupación prolongada puede haber sido la utilización de la madera como combustible. Los experimentos han demostrado que es necesaria alguna madera para utilizar el hueso como combustible. Esta población entró en las Américas a través de una ruta costera hace ca 16,0 ka, cuando quedó disponible un camino viable (Lesnek et al, 2018).
De acuerdo con evidencias genéticas, la población fundadora fue de unos pocos cientos de individuos (Hey, 2005; Fagundes et al, 2007; Kitchen et al, 2008; Gutenkunst et al, 2009; Ray et al, 2010; Wall et al, 2011; Gravel et al, 2013; Fagundes et al, 2018).
 |
| Modelo elaborado por la Universidad de Cambridge a partir de Moreno-Mayar et al, 2018. |
El tiempo en alcanzar el Sur del continente fue de ca 1,4 ka, ya que Monte Verde, en el sur de Chile, se ha datado en ca 14,6 ka.
- Siguieron la costa occidental, por caminos hoy bajo el nivel del mar, lo que explica que no hayan aparecido restos. Esta opción permitiría la pesca. Desde hace 14 ka este pasillo estuvo exento de hielo y podía encontrarse megafauna. Esta opción ha sido defendida por Knut Fladmark y por Dixon (1999, 2001).
- La teoría de la migración costera sostiene el desplazamiento por mar de los primeros colonos, utilizando embarcaciones para desplazarse cerca de la costa aprovechando la disponibilidad de recursos acuáticos abundantes, alimentados por bosques de algas (Erlandson et al, 2007; Braje et al, 2017; kelp highway hypothesis). Las islas Ryukyu fueron pobladas hace 35 ka.
- Algunos investigadores defienden una ruta a través del interior del continente, por el territorio de Yukon y Alberta. Heintzman et al (2016) y Pedersen et al (2016) encontraron evidencia de un corredor para el paso del bisonte hace 13 ka. Hace ca 11,5-11 ka se abrió una ruta a través de un corredor libre de hielo (ice-free corridor) a lo largo del lado oriental de las Montañas Rocosas. Hay muchos yacimientos fechados entre 11,5-11 ka y que se caracterizan por la presencia de puntas de flecha de la cultura Clovis (Modo 4), que toma su nombre del yacimiento de Nuevo México donde fue definida por primera vez.
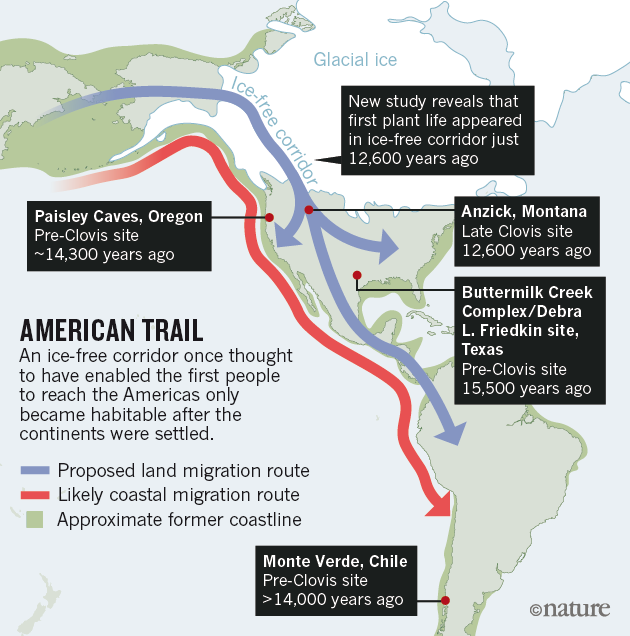 |
| Rutas de entrada de los humanos en América |
- Para unos investigadores, en tres grandes grupos:
- los esquimales-aleutianos, quienes se instalaron en las islas Aleutianas, en Alaska y en la costa norte de Norteamérica.
- Na Dene, llamados así por la familia de lenguajes que lleva ese nombre, que se asentaron en Alaska y en la costa noroeste de Norteamérica hasta el comienzo del segundo milenio de nuestra era, momento en que sus descendientes emigraron más al sur y al este hasta los territorios que hoy forman Canadá y los Estados Unidos. Sus representantes más meridionales son los apaches y los navajo que se establecieron en el sudoeste de América del Norte hacia el año 1200 a.C.
- Amerindios.
- Para otros, en cientos de grupos independientes Paleo-indios, en varias olas migratorias.
Se ha sugerido una conexión entre las lenguas del Yenisei y las lenguas Na-Dene. Aceptando esta relación, Mark A. Sicoli y Gary Holton (2014) han utilizado datos lingüísticos tipológicos (gramaticales) y modelos filogenéticos computacionales. Los resultados apoyan una dispersión temprana de Na-Dene desde Beringia, a lo largo de la costa de América del Norte, con una migración de vuelta de los Yeniseicos a través de Siberia y una propagación posterior de lenguas Na-Dene por el interior de América del Norte.
Este escenario se basa en el razonamiento geográfico y en los datos genéticos (Llamas et al, 2016; Lindo et al (2017); pero los patrones observados pueden deberse también a la deriva genética) y lingüísticos, pero no ha sido confirmado por las pruebas arqueológicas.
Se han propuesto otros lugares alternativos, como la isla japonesa de Hokkaido y la isla rusa de Sajalín. Durante el máximo glacial, estas islas, junto con las Kuriles, estaban conectadas y hay signos de presencia humana hace 26 ka.
Se han propuesto otros lugares alternativos, como la isla japonesa de Hokkaido y la isla rusa de Sajalín. Durante el máximo glacial, estas islas, junto con las Kuriles, estaban conectadas y hay signos de presencia humana hace 26 ka.
La Teoría de las Tres Migraciones
- Sundadontia. En pueblos del sureste de Asia y en poblaciones prehistóricas de América. Se trata de una dentición menos especializada.
- Sinodontia. Este patrón está presente en los nativos americanos y del noreste de Asia. Dentro de este patrón identificó tres grandes subdivisiones que corresponderían a tres migraciones distintas.
- Este patrón pudo haber evolucionado desde la sundadontia por aislamiento genético hace 20 ka (Stojanowski et al, 2013)
- Dos migraciones costeras, una por el Pacífico (norte y sur de América) y otra por el Atlántico (norte de América).
- Una migración posterior a través de un corredor libre de hielo hacia el centro de Norteamérica.
Oleadas migratorias
Las poblaciones nativas de América del Sur presentan una diversidad craneométrica muy alta. Junto con los estudios genéticos, estos datos evidencian que América fue poblada por varias oleadas de dispersión procedentes del noreste de Asia a lo largo del Pleistoceno y a principios del Holoceno (Noreen von Cramon Taubadel, André Strauss y Marcos Hubbe, 2017).| Algunas herramientas de Cooper's Ferry utilizadas para la caza y la pesca. Davis et al (2019). |
Migración costera desde Japón hace 16 ka.
Partiendo del hallazo de herramientas líticas en Cooper's Ferry, Davis et al (2019) plantean la posibilidad de una migración desde Japón a través de una ruta por la costa del Pacífico.
- La datación de los hallazgos (hace 16,56-15,28 ka) es anterior a la apertura de rutas desde Beringia.
- Los artefactos son similares a los más antiguos recuperados en la isla de Hokkaido.
Evidencias genéticas
Los primeros estudios genéticos analizaron en ADNmt, pero en la última década, los avances tecnológicos han permitido la reconstrucción de los genomas prehistóricos. Estos estudios genómicos han demostrado de manera concluyente que los primeros estadounidenses fueron el resultado de una mezcla ancestral de Asia Oriental y el Norte de Eurasia. Esta población fundadora se abrió camino hacia el este de Beringia y tras una división de poblaciones, viajó al sur de las capas de hielo continentales que cubrían Canadá en algún momento entre hace ca 17,5-14,6 ka. Estos resultados genéticos concuerdan con el registro arqueológico (Waters et al, 2019).Genoma completo de USR1
Moreno-Mayar et al (2018) obtuvieron el genoma completo de USR1. De acuerdo con los resultados, este individuo perteneció a una población diferente a la de los nativos americanos actuales y antiguos cuyo genoma se ha secuenciado hasta la fecha.
- Usando modelos demográficos, han inferido que todas las poblaciones nativas americanas descienden de una población fundadora que comenzó a separarse de los asiáticos del este hace 36 ± 1,5 ka. Hace 25 ka, el intercambio genético se detuvo.
- He Yu et al (2020) han hallado en un individuo de hace 14 ka de la región del Lago Baikal y en otro del Mesolítico del noreste de Siberia una mezcla genética compartida con individuos de Eurasia del Norte y nativos americanos no árticos, lo que sugiere que la ascendencia que más tarde dio lugar a los nativos americanos en América del Norte y del Sur estaba mucho más ampliamente distribuida de lo que se suponía.
- Hace 20 ka el grupo de USR1 se separó de las poblaciones ancestrales de los nativos americanos modernos.
- Por otra parte, depués de hace 11,5 ka, algunas poblaciones americanas hibridaron con una población siberiana cercana a los Koryaks.
- Las ramas basales de los nativos americanos del norte y del sur comenzaron a diverger hace ca 17,5–14,6 ka.
- Estas conclusiones apoyan la hipótesis Standstill of Beringia.
 |
Representación esquemática de los procesos de dispersión humana y divergencia en las Américas, ordenados cronológicamente (Moreno-Mayar et al, 2018).
|
Varios genomas antiguos
Raghavan et al (2015) analizaron 31 genomas modernos de América, Siberia y Oceanía; y 23 genomas antiguos de toda América (de hace 6000-200 años). Los autores concluyeron que todos los nativos americanos forman un grupo monofilético que se dividió en dos ramas: amerindios y atabascas. La fecha más probable para esta separación es la de hace 22 ka, aunque parece que el flujo genético continuó hasta hace unos 12 ka, momento que coincide con el aumento del nivel del mar y la desaparición bajo las aguas del puente de Beringia.
En relación a un conjunto de datos de poblaciones actuales, y los genomas antiguos disponibles de Anzick1, Montana (hace ca 12,8 ka), Kennewick Man/Ancient One, Washington (ca 9 ka) y USR1, Alaska (ca 11,5 ka), Moreno Mayar et al (2018) han obtenido y analizado los siguientes genomas:
- Trail Creek Cave 2, Alaska (datado en hace ca 9 ka).
- Big Bar Lake, British Columbia ca 5,6 ka.
- Spirit Cave, Nevada ca 10,7 ka.
- Cuatro individuos de Lovelock Cave, Nevada (ca 1,95-0,6 ka)
- Cinco individuos de Lagoa Santa, Brasil (ca 10,4-9,8 ka)
- Dos indiviuos de Punta Santa Ana y Ayayema en Patagonia, Chile (ca 7,2-5,1 ka)
- Momia inca de Mendoza, Argentina, ca 0,5 ka.
- Individuo del XIX de las Islas Andamán.
De acuerdo con las conclusiones:
- De los antiguos beringios (USR1 y Trail Creek), se desprendieron dos grupos, en el sureste de Beringia (Alaska y Yukón), hace ca 17,5-14,6 ka.
- Nativos Americanos del Norte. Los atabascanos e inuit, que habitan actualmente en Alaska, se trasladaron hacia el norte en algún momento después de hace 9 ka, la edad del individuo de Trail Creek.
- Nativos Americanos del Sur (Spirit Cave), con dispersión por las Rocosas.
- Un grupo, que se separó hace unos 14,1 ka, alcanzó América del Sur rápidamente.
- Los Mesoamericanos se separaron de los sudamericanos poco después de hace 13,9 ka.
- Los datos de Spirit Cave y Lagoa Santa indican que el patrón de dispersión por América del Sur incluye eventos complejos de mezcla de poblaciones, a lo largo de múltiples caminos geográficos.
- Los suruí amazónicos y Lagoa Santa comparten una mayor proporción de alelos con grupos de Australasia (representados por papuanos, australianos e isleños de Andaman), que los Mesoamericanos.
- Esta señal no ha sido documentada en América del Norte, lo que implica que el grupo que la poseía ha desaparecido o que una nueva población llegó más tarde y pasó por Norteamérica sin dejar traza genética.
- La mayoría de las poblaciones nativas actuales de América del Sur, forman un clado con Lagoa Santa, pero derivan de una mezcla con ancestros mesoamericanos que llegaron en una segunda migración independiente, en mayor grado al oeste de los Andes.
- El genoma Ayayema no muestra esta relación con Mesoamérica, lo que sugiere que estas poblaciones llegaron a América del Sur después de hace 5,1 ka o que no alcanzaron la remota región patagónica.
- La historia de los habitantes del noreste de América estuvo marcada por una mezcla entre lo antecesores de los actuales nativos americanos del norte y del sur y por el aislamiento entre grupos en la zona costera.
- Las poblaciones se expandieron por América del Norte tal vez en unas pocas centenas de años y por América del Sur en mil o dos mil años. Tras los asentamientos iniciales, se produjeron eventos complejos de movimiento y mezcla de poblaciones. Desde el sureste de Beringia, los nativos americanos se irradiaron rápidamente, no de forma gradual, sino a saltos, y dieron origen a múltiples poblaciones que en diferentes momentos se expandieron a diferentes partes del continente, aunque no tan extensivamente como la población inicial.
Cromosoma Y
Se pueden observar dos linajes fundadores de ascendencia asiática:
- Q, representa el 75% de los ADN-Y de los nativos americanos. Se observa por todo el continente.
- Según Vincenza Battaglia et al (2013) se distinguen dos linajes fundadores principales, Q1a3a1a-M3 y Q1a3a1-L54 (XM3), junto con sub-clados de menor edad y distribución geográfica más restringida.
- Q1a3a1a-M3 se observa también en el Lejano Oriente asiático.
- Q1a3a1-L54 (XM3) se encuentra en el sur de Siberia, representado por el sub-clado Q1a3a1c-L330.
- Estos datos confirman un origen siberiano sur de las poblaciones ancestrales que dieron origen a los Paleo-indios y la diferenciación de los dos Q linajes fundadores nativos en Beringia.
- Los dos linajes fundadores llegaron a Mesoamérica al mismo tiempo y desde allí iniciaron una rápida migración hacia el sur, a lo largo de la costa del Pacífico, en la región andina.
- Los dos linajes, muestran diferentes patrones de evolución en el altiplano mexicano y el área andina, lo que puede explicarse por las diferenciaciones locales debido a los eventos demográficos provocados por la introducción de la agricultura y asociados con el florecimiento de los grandes imperios.
- El ADN-Y linaje de Anzick es Q1a2a1 * (L54). Entre las modernas secuencias comparadas, la de un Maya es la más cercana.
- C, representa el 6% de los ADN-Y de los nativos americanos. Se limita a América del Norte. Es probable que su llegada fuese posterior (Lell et al, 2002; Pero Zegura et al, 2004).
- El linaje más antiguo C3* sólo se ha detectado significativamente en un área restringida de Ecuador (Kichwa y Waorani, culturalmente muy diferenciados, conocidos los últimos por su extrema ferocidad con los extraños, genéticamente aislados entre ellos) Este haplotipo es prácticamente ausente en América del Norte y Central, pero se encuentra en el centro, este y noreste de Asia con una frecuencia elevada. Esto sugiere una introducción tardía (más de 6 ka) a través de rutas costeras o cruzando el Pacífico y que las poblaciones que migraron fueron muy reducidas. Dada la similitud de los artefactos de cerámica encontrados en ambas regiones, se ha especulado con una conexión a través del Pacífico entre la cultura Jomon medio de Kyushu (Japón) y la cultura Valdivia de Ecuador en 4400-3300 aC (Lutz Roewer et al, 2013).
ADN-mt
Los linajes indígenas americanos comienzan con las letras A, B, C, D o X.
Tamm et al (2007) analizaron el mitogenoma de 601 individuos de América (pertenecientes a 20 poblaciones de todo el continente) junto a 3764 muestras de Asia pertenecientes a 26 poblaciones diferentes. Los resultados revelaron que los americanos poseían tres variaciones únicas –identificadas como subclados «C1b», «C1c» y «C1d»– ampliamente distribuidas por el continente americano. La cronología basada en las tasas de mutación, les llevó a concluir que los antepasados de los primeros americanos quedaron aislados de sus parientes asiáticos hace unos 25 ka, antes de que entraran definitivamente en América hace unos 15ka para distribuirse rápidamente por todo el continente. Un análisis de la distribución geográfica de estos marcadores genéticos apuntó a que el lugar más probable donde se pudo producir ese aislamiento fue Beringia.
Tamm et al (2007) analizaron el mitogenoma de 601 individuos de América (pertenecientes a 20 poblaciones de todo el continente) junto a 3764 muestras de Asia pertenecientes a 26 poblaciones diferentes. Los resultados revelaron que los americanos poseían tres variaciones únicas –identificadas como subclados «C1b», «C1c» y «C1d»– ampliamente distribuidas por el continente americano. La cronología basada en las tasas de mutación, les llevó a concluir que los antepasados de los primeros americanos quedaron aislados de sus parientes asiáticos hace unos 25 ka, antes de que entraran definitivamente en América hace unos 15ka para distribuirse rápidamente por todo el continente. Un análisis de la distribución geográfica de estos marcadores genéticos apuntó a que el lugar más probable donde se pudo producir ese aislamiento fue Beringia.
- Hace 18-15 ka, la primera ola migratoria llegó desde Asia hasta la costa del Pacífico y desde allí se movió hacia el interior. Esa primera ola migratoria, con el haplogrupo B2, contenía los ancestros de todas las tribus de Norteamérica, Centroamérica y América del Sur.
- El haplogrupo B2a surgió en América del Norte hace 13-12 ka. Está ausente en los esquimo-aleutianos y en los hablantes Na-Dene del norte.
- Una segunda ola de migración probablemente dejó Siberia sólo un par de miles de años después de la primera ola. En lugar de recorrer la costa, el segundo grupo se deslizó a través de un corredor libre de hielo que va desde Alaska al sur de Canadá. Esta segunda ola nunca llegó al sur de Estados Unidos. Los recién llegados y las poblaciones existentes hibridaron. Las marcas genéticas de esta ola (haplogrupos X2a y C4c), se pueden observar tanto en hablantes Na-Dene como no-Na-Dene.
- Una tercera ola de migración, llevada a cabo por descendientes de los pobladores ancestrales, comenzó alrededor de hace 7-4 ka en Alaska y se extendió hacia el oeste de Asia y sobre todo hacia el este del Canadá circumpolar. El haplogrupo (A2a) es típico de esquimo-aleutinanos y Na-Dene y también está presente en los siberianos orientales.
Naia, la chica de Hoyo Negro, presenta el haplogrupo D1, que se cree procede de Beringia, y es exclusivo actualmente de los nativos americanos incluyendo sudamérica.
Los bebés de Upward Sun River, presentan los haplogrupos C1b y B2, presentes pero raros en los americanos actuales pero ausentes en Asia (Tackney et al, 2015)
- C1 se encuentra con mayor frecuencia entre los indios Pima y Hualapai de Arizona, Delta Yuman de California, y otras seis tribus, incluyendo la Ignaciano en Bolivia, los extintos taínos de Puerto Rico y un grupo representado por los huesos de Norris Farms en Illinois, datados en hace 700 años.
- B2 se encuentra con mayor frecuencia en 37 tribus a lo largo de las Américas, incluyendo los Yakama, Wishram, Northern Paiute-Shoshone, Navajo, Hualapai (donde también aparece C1), Zuni y Jemez en América del Norte y Quechua y Aymara en Perú. También era común en los antigos Fremont y Anasazi del suroeste de USA.
Bastien Llamas et al (2016) han secuenciado el ADN-mt completo de 92 individuos precolombinos datados entre 8,6-0,5 ka para reconstruir de forma detallada la historia de la población de las Américas mediante un análisis de coalescencia por inferencia bayesiana. Los datos construidos sugieren que una pequeña población entró en las Américas a través de una ruta costera alrededor de ca 16,0 ka, después de un aislamiento previo en Beringia de ca 2,4-9 ka. Todos los antiguos linajes mitocondriales detectados en este estudio están ausentes de los conjuntos de datos modernos, lo que sugiere una alta tasa de extinción. La colonización europea provocó una pérdida sustancial de linajes precolombinos.
- Las 92 muestras corresponden a 84 haplotipos distintos, dentro de los haplogrupos A2, B2, C1b, C1c, C1d, y D1. Ninguno de estos 84 haplotipos están representados en la población humana actual.
- Todos los haplotipos formaban parte de una población inicial. Cada haplotipo parece estar limitado a una región geográfica concreta o a unos antecedentes culturales compartidos, lo cual es consistente con la sugerencia de que la estructura geográfica se estableció rápidamente y el flujo de genes posterior fue limitado.
- El linaje compartido entre la población siberiana y la ancestral americana data de hace 24,9 ka. Las dos poblaciones quedaron totalmente aisladas entre hace 24,9-18,4 ka, coincidiendo con el Último Glacial Máximo (LGM), que pudo obligar a las poblaciones siberianas a buscar refugios más al sur mientras que otros grupos permanecían aislados en zonas libres de hielo en Beringia ante la imposibilidad de retirarse más al sur. La población femenina eficaz de la población fundadora que posteriormente entró en América era de ca 2.000 individuos. La megafauna de Beringia fue más que suficiente para sostener a esta población.
- El arbol mitogenómico muestra un repentino estallido de diversificación de linajes hace ca 16-13 ka, seguido por un marcado aumento en el tamaño efectivo de la población femenina. En general, la población se multiplicó por 60 entre hace 16-13 ka, lo que sugiere un entorno más favorable, ya en América, con un rápido retroceso de los glaciares costeros. La estancia en Beringia duró por consiguiente 2,4-9 ka y el tiempo en alcanzar el Sur del continente fue de ca 1,4 ka, ya que Monte Verde, en el sur de Chile, se ha datado en ca 14,6 ka.
- El ancestro común más reciente de las muestras antiguas y los nativos actuales data de hace ca 9 ka. Las altas tasas de extinción después de la colonización europea en poblaciones separadas geográficamente fueron la probable causa de la desaparición de la diversidad genética en ciertas áreas.
Lindo et al (2017) secuenciarion el genoma de “Shuká K áa”, recuperado en On Your Knees Cave (OYKC) en el sureste de Alaska (sitio arqueológico 49-PET-408) y datado en hace ca 10,3 ka cal, y de tres individuos de la Columbia Británica datados hace ca 6.075-1.750 años cal. De acuerdo con los resultados, la costa del noroeste del Pacífico exhibe continuidad genética durante al menos los últimos 10.300 años. “Shuká K áa” pertenece a una línea ancestral diferente a Anzick-1 y Kennewick Man). Los individuos analizados de la costa noroeste pertenecen a un linaje genético precoz que puede provenir de una migración costera de finales del Pleistoceno.
 |
| La migraciones americanas a la luz del ADN-mt. Amick, 2016. |
Complejo HLA
Según A. Arnaiz-Villena et al (2006), del análisis de los genes HLA, se infiere que los Amerindios son los primeros habitantes de las Américas y estaban allí cuando llegaron oleadas de gentes hablando lenguas Na Dene (Atabascos, Navajos, Apaches) y Esquimales. Mientras se observa que en todas las otras poblaciones mundiales existe un gradiente de emparentamiento, que va en general concordando con la proximidad o lejanía geográfica, los Amerindios se sitúan aparte de todos.- Los atabascos del noroeste canadiense muestran flujo génico con poblaciones vecinas, con amerindios, con habitantes de las islas del Pacífico, incluyendo australianos orientales, y con siberianos, ya que comparten haplotipos DRB1-DQB1 con estas poblaciones (por ejemplo: DRB1*14:01-DQB1*05:03, DRB1*09:01-DQB1*03:03).
- La entrada de los amerindios en América pudo haber sido diferente a la de atabascos, aleutianos y esquimales; los amerindios pudieron haber llegado al continente mucho antes que los atabascos y esquimales ya que presentan un conjunto completamente diferente de frecuencias alélicas HLA-DRB1.
- Los amerindios muestran muy pocos alelos estrictamente particulares (DRB1*04:11, DRB1*04:17), pero presentan haplotipos extendidos únicos (por ejemplo: A*02-B*35-DRB1*04:07-DQB1*03:02,A*02-B*35-DRB1*08:02-DQB1*04:02).
- Estos resultados no apoyan el modelo clásico de poblamiento del continente de las tres oleadas migratorias, sino otro en el que la entrada pudo ser también por la costa pacífica. La llegada de gentes por el Océano Pacífico ha podido contribuir al perfil genético HLA americano. La migración inversa (de América a Asia) de gentes en diferentes épocas no se puede descartar.
Grupos sanguíneos
Fernando A. Villanea et al (2013) han estudiado la distribución geográfica y temporal de una variante única del grupo sanguíneo O llamado O1vG542A, común entre los nativos meso y sudamericanos, pero poco frecuente en otras poblaciones.
Identificaron este alelo en una población Na-Dene a una frecuencia de 0,11, y una antigua población de California a una frecuencia de 0,20. Estos resultados apoyan un origen en Beringia de O1vG542A. Este resultado es consistente con la hipótesis de que las poblaciones de nativos americanos derivan principalmente su ascendencia a partir de una única población de origen (out of Beringia).
 |
| Árbol genético de máxima probabilidad. |
SNPs
Reich et al (2012) encontraron evidencia de al menos tres eventos de flujo de genes desde Asia hasta el Nuevo Mundo, asociados con grupos na-dene, esquimales-aleutianos y amerindios. La ascendencia amerindia divergió primero, mientras que la ascendencia na-dene y eskimo-aleutiana proviene de una población de origen siberiano ancestral común. El análisis también identifica una migración de vuelta de América del Norte a Siberia ancestral en los antepasados de los Naukan Yupik, que posteriormente se mezclaron con las poblaciones Chukchi.
Amorim et al (2017) han hallado señales de selección natural positiva en las desaturasas de ácidos grasos (FADS), unos genes que hacen posible la digestión de alimentos ricos en grasas insaturadas, en todas las poblaciones nativas americanas lo que sugiere un evento de adaptación única y fuerte que se produjo en Beringia, antes de la expansión por América.
 |
Historia de las poblaciones nativas americanas, inferida a partir del genoma.
Los círculos de color indican poblaciones ancestrales inferidas.
Las líneas continuas, descendencia. Los números indican la deriva genética (en unidades proporcionales a la distancia genética).
Las líneas de puntos indican eventos de mezcla, con el porcentaje de asociado a la contribución.
El color rojo indica grupos Eskimo-Aleutianos.
El verde, grupos Na-Dene.
El azul, grupos amerindios.
|
Flujo genético de antiguas poblaciones siberianas
Maanasa Raghavan et al (2013) secuenciaron el ADN de un individuo juvenil (MA 1) cuyos restos hallados en Mal'ta (Lago Baikal, centro-sur de Siberia), fueron datados en 24 ka. Se trata del genoma humano anatómicamente moderno más antiguo reportado hasta la fecha.
- El ADN-mt pertenece al haplogrupo U, con alta frecuencia entre los cazadores recolectores del Paleolítico Superior y Mesolítico europeo.
- El ADN-Y pertenece al haplogrupo R, basal a euroasiáticos occidentales de hoy en día y es cercano a la raíz de la mayoría de los linajes nativos americanos.
- El ADN autosómico es basal a euroasiáticos occidentales de hoy en día y genéticamente muy relacionado con los actuales nativos americanos, sin afinidad con los asiáticos orientales.
La presencia de genes de la población a la que pertenecía MA-1 en el linaje de los nativos americanos podría explicar por qué algunos cráneos de los primeros americanos muestran características morfológicas que no se parecen a las de los asiáticos orientales.
Hipótesis descartadas
- El modelo tradicional (blitzkrieg) fue propuesto por Hrdlička y posteriormente modificado con el fin de incorporar los nuevos datos cronológicos y culturales. Supone una rápida migración (que contiene todos los linajes fundadores) de ancestros asiáticos a través de de Bering hace unos 16,5 ka. La base de este modelo fue la identificación de la cultura Clovis.
- Greenberg et al, combinando pruebas lingüísticas, arqueológicas y biológicas, propusieron un modelo de tres oleadas migratorias, según el cual los tres grupos lingüísticos nativos americanos propuestos (amerindios, Na-Dene y Eskimo-Aleut) entraron separadamente en las Américas después del último máximo glacial (LGM, ~ 20 kya).
- Varias docenas de herramientas de tecnología solutrense, datadas entre 26-19 ka, han sido descubiertas en seis lugares a lo largo de la costa este de Norteamérica. Dennis Stanford y Bruce Bradley (2012) han propuesto que gentes de Europa occidental migraron a América del Norte desplazándose (por encima de la superficie del hielo y/o en barcas) a lo largo del borde de la parte congelada del Atlántico Norte. La zona de cambio estacional, donde el hielo finalizaba y comenzaba el océano abierto, habría sido extremadamente rica en recursos alimenticios. Por otra parte, la presencia del haplogrupo X2a en el ADN-mt de los nativos americanos ha dado pie a hipótesis de un flujo genético transatlántico, que según Raff y Bolnick (2015) son inconsistentes (ir a comentarios de José Luis Moreno Garvayo; y aquí).
 |
| Yacimientos arqueológicos tempranos a lo largo de la vertiente andina occidental entre 16 ° y 25 ° S. Puntos oscuros: sitios costeros, puntos verdes: sitios ubicados en altitudes superiores a 3.000 m snm. La zona amarilla representa la extensión del núcleo del desierto de Atacama. |
Este comentario ha sido eliminado por un administrador del blog.
ResponderEliminarThanks for sharing.
ResponderEliminarRe puto
ResponderEliminarHijos puta
ResponderEliminarNo es por nada pero el gregory le gusta los hombres mmm...
ResponderEliminar