 |
| Resumen de especies domesticadas a lo largo del tiempo. Greger Larson y Dorian P. Fuller (2014) |
Greger Larson y Dorian P. Fuller (2014) resumen los caminos evolutivos seguidos en la domesticación de los animales.
- Comensalismo (Commensal Pathway) o autodomesticación.
- No existe una intención humana.
- Diferentes poblaciones de animales salvajes habrían sido atraídas por elementos humanos, incluidos los residuos de alimentos y / o animales más pequeños atraídos por estos residuos. Estas poblaciones sinántropas estarían formadas por individuos más dóciles y menos agresivos.
- En el caso del perro, Coppinger y Coppinger plantearon la hipótesis de que la domesticación del perro se produjo durante el Neolítico cuando asentamientos humanos proporcionaron un nuevo nicho debido a la disponibilidad permanente de residuos de alimentos. El experimento de Belyaev con zorros plateados (Vulpes vulpes) mostró cómo la domesticación puede tener lugar rápidamente cuando una fuente de alimento está fácilmente disponible. El establecimiento de asentamientos permanentes en el Neolítico habría creado un ambiente donde la selección de la mansedumbre pudo mantenerse durante muchas generaciones estableciendo así las bases para la domesticación del perro.
- Para Zeder (2012) el paso de una población sinantrópica a una domesticada sólo podría haber tenido lugar después de que los animales progresasen desde la antropofilia a la habituación, al comensalismo y la asociación, momento en el que el establecimiento de una relación recíproca entre animales y humanos habría sentado las bases para la domesticación, incluyendo el cautiverio y la reproducción humana controlada.
- Desde esta perspectiva, la domesticación de animales es un proceso de coevolución axiomático en el que una población responde a la presión selectiva adaptándose a un nuevo nicho que incluye otras especies con comportamientos cambiantes.
- Los mecanismos que redujeron el flujo de genes entre las poblaciones salvajes y las emergentes puede que no hayan sido autoevidentes. Fiona Marshall et al (2014) tras revisar las investigaciones recientes sobre la domesticación de burros, camélidos (que incluye los dromedarios, camellos bactrianos, llamas y alpacas) cerdos, vacas, ovejas y cabras sugiere que ni la cría intencional ni el aislamiento genético eran tan significativos como se pensaba tradicionalmente. Por el contrario, los resultados muestran poco control de la reproducción e indican el flujo de genes con animales salvajes.
- Los perros son el ejemplo típico de este tipo de domesticación. Se trata del único animal domesticado antes del advenimiento de la agricultura.
- Adam H. Freedman et al (2013) analizaron el genoma de tres lobos grises, dos perros de linajes antiguos y un chacal. Los perros y los lobos divergieron por un proceso dinámico, con cuellos de botella de población en ambos linajes y flujo génico post-divergencia.
- Los lobos grises se extendían por todo el hemisferio norte y por ello es difícil establecer donde fueron domesticados o incluso si el proceso se desarrolló varias veces en múltiples lugares de forma independiente. Los lobos competían con los humanos por los mismos recursos; teniendo en cuenta que las poblaciones humanas eran cazadoras-recolectoras móviles, no es obvio cómo surgieron los perros y cómo se modificó su naturaleza salvaje. Es posible que los lobos con antropofilia formasen un ecotipo sinántropo diferente al de los demás lobos que conservase su integridad aun en el caso de flujo genético y que hubiese desarrollado adaptaciones específicas al nicho humano. Cuando la diferencia fue apreciable, quizá se produjo la domesticación gracias a una selección humana más intensiva. Actualmente, poblaciones diferenciadas de lobos, zorros y ballenas que comparten territorios pero explotan recursos distintos, muestran diferentes
adaptaciones fenotípicas y de comportamiento. - Otros: gatos, palomas, cerdos de guinea, ratas polinesias, quizá el cerdo en China, gallina,
- Domesticación de presas de caza (Prey Pathway)
- Existe intención humana: aumentar la eficiencia en la gestión de los recursos alimenticios aumentando la disponibilidad de grandes y medianos herbívoros. La domesticación es un resultado añadido.
- La selección de rasgos como la docilidad permitió un control más completo sobre la dieta y la reproducción de los animales (Zeder, 2012).
- Está asociada con las comunidades agrícolas. Marom y Bar-Oz (2013) han publicado un estudio de la interacción humana con los bóvidos y los suidos (Bos taurus y Sus scrofa) en el Valle del Jordán. La incorporación de bóvidos y suidos sucedió hace 8 ka. La sobrecaza y disminución de las poblaciones salvajes, precedió a la domesticación. La selección natural actuó sobre estas especies con los siguientes efectos:
- Disminución de tamaño.
- Aumento de la tasa de rotación, con una edad más joven para la procreación.
- Neotenia.
- Además, la eliminación de los individuos más grandes y agresivos, permitió a los humanos tomar su papel como líderes en la estructura jerárquica.
- En muchos casos se produjo una diversificación secundaria para desarrollar cualidades diferentes a la obtención de carne, incluyendo fibras textiles, leche y tracción.
- Zohary et al (1998), consideran que la sobrecaza que la precedió es indicio de falta de intencionalidad de la domesticación.
- Siguieron este camino la domesticación de cabras, ovejas, vacas, llamas, alpacas, pavos y patos.
- El reno es otro ejemplo, con la particularidad de que fue domesticado por sociedades de cazadores.
- Domesticación intencional (Directed Pathway) o domesticación social.
- Tras haber conseguido la domesticación de las primeras plantas y animales, los humanos lo intentaron con nuevas especies como caballos, burros, camellos y dromedarios.
- La domesticación se produce mediante la selección activa de las crías más dóciles y no se logró cuando no fue posible conseguir la docilidad (gacelas o cebras).
- Otros casos: abejas, gusanos de seda, conejos, pájaros y peces.
 |
| Lugar y tipo de domesticación. Greger Larson y Dorian P. Fuller (2014) |
PERROS
Pat Shipman (2014) explica desde el punto de vista tafonómico la presencia de yacimientos en Eurasia, datados entre 40-15 ka (tras la aparición del HAM en Eurasia), con una cantidad inusualmente grande de restos de mamut, abundantes artefactos y, a menudo, viviendas construidas con huesos de mamut.- El gran número de individuos y la escasez de marcas de dientes de carnívoros sugieren una nueva capacidad de retención de las presas.
- Los patrones de edad de las presas en algunos yacimientos se asemejan con independencia de las distancias temporales y espaciales de las localizaciones, lo que sugiere una transmisión tecnológica o de comportamiento.
- La hipótesis del autor es que estas innovaciones pudieron haber sido facilitadas por un temprano intento de domesticación de los perros.
En su libro The Invaders (2015) Shipman propone que los perros ayudaron a los HAM a extinguir a los neandertales.
Evidencias arqueológicas
El hallazgo con datación más antigua, corresponde a la Cueva Razboinichya en los montes Altai (Siberia) donde se localizó un cráneo, una mandíbula y varios dientes en perfecto estado de conservación, datados en 33 ka.En el yacimiento belga de Goyet durante de una excavación en el siglo XIX, aparecieron restos datados en 31,7 ka. Presenta una mayor capacidad craneal y un hocico más corto y ancho, pero sobretodo un mayor tamaño en los dientes. Un estudio de Abby Grace Drake, Michael Coquerelle y Guillaume Colombeau (2015) demostró que corresponden a un lobo.
En el yacimiento gravetiense de Předmostí, Moravia, República Checa, se encontraron restos de primitivos perros (2011), datados en 26-24 ka. Uno de los cráneos presenta especial interés ya que fue enterrado ritualmente con un hueso de mamut entre los dientes. Los investigadores plantearon en su momento la hipótesis de que fueron empleados como animales de carga debido a su gran tamaño. A partir de los isótopos en los huesos, Bocherens et al (2014) consideran que fueron alimentados con reno y buey almizclero y esta dieta apoyaría la hipótesis del control humano de su dieta y del uso para transporte, ya que la caza estaba basada en el mamut.
La cueva de Chauvet ha conservado las huellas de un niño acompañado por un perro. Restos de antorcha han permitido datar las pisadas mediante el análisis del C14 en más de 26 ka.
En Eliseevichi I, en la cuenca del río Dniéper, en las
estepas de Ucrania, se encontraron evidencias de perros datadas entre
17-13 ka.
El perro de Mezhirich (Ucrania) está datado en 15 ka y el de Kesserloch (Suiza) en 14-12 uncal ka.
El perro de Mezhirich (Ucrania) está datado en 15 ka y el de Kesserloch (Suiza) en 14-12 uncal ka.
Esto quiere decir que la transformación de lobo a perro ocurrió
mucho antes y que el lobo fue el primer animal domesticado por el sapiens.
En Bonn-Oberkassel, con una datación de hace 14223 años, se ha reportado un enterramiento de dos perros junto con dos humanos adultos y ajuares. Uno de los perros era un juvenil tardío cuando fue enterrado. Las lesiones de la cavidad oral indican un perro gravemente enfermo que probablemente sufrió una infección por morbilivirus (moquillo canino). Aparece una línea dental de sugestiva hipoplasia del esmalte en la etapa de desarrollo de 19 semanas. Dos líneas adicionales de hipoplasia del esmalte, solo en el canino, documentan más episodios de enfermedad en las semanas 21 y 23. Los cambios patológicos también incluyen enfermedad periodontal grave que puede haber sido facilitada por la inmunodeficiencia. Debido a que el moquillo canino tiene un curso de enfermedad de tres semanas con una mortalidad muy alta, el perro debe haber estado enfermo perniciosamente durante los tres episodios de enfermedad y entre las edades de 19 y 23 semanas. La supervivencia sin asistencia humana intensiva hubiera sido poco probable. Antes y durante este período, el perro no puede haber tenido ningún uso utilitario para los humanos. Probablemente algunos humanos del Pleistoceno tardío consideraron a los perros no solo materialmente, sino que también pudieron haber desarrollado vínculos emocionales y afectivos con sus perros, tal como lo refleja la supervivencia de este perro, posiblemente a través del cuidado humano (Janssens et al, 2018)
En Bonn-Oberkassel, con una datación de hace 14223 años, se ha reportado un enterramiento de dos perros junto con dos humanos adultos y ajuares. Uno de los perros era un juvenil tardío cuando fue enterrado. Las lesiones de la cavidad oral indican un perro gravemente enfermo que probablemente sufrió una infección por morbilivirus (moquillo canino). Aparece una línea dental de sugestiva hipoplasia del esmalte en la etapa de desarrollo de 19 semanas. Dos líneas adicionales de hipoplasia del esmalte, solo en el canino, documentan más episodios de enfermedad en las semanas 21 y 23. Los cambios patológicos también incluyen enfermedad periodontal grave que puede haber sido facilitada por la inmunodeficiencia. Debido a que el moquillo canino tiene un curso de enfermedad de tres semanas con una mortalidad muy alta, el perro debe haber estado enfermo perniciosamente durante los tres episodios de enfermedad y entre las edades de 19 y 23 semanas. La supervivencia sin asistencia humana intensiva hubiera sido poco probable. Antes y durante este período, el perro no puede haber tenido ningún uso utilitario para los humanos. Probablemente algunos humanos del Pleistoceno tardío consideraron a los perros no solo materialmente, sino que también pudieron haber desarrollado vínculos emocionales y afectivos con sus perros, tal como lo refleja la supervivencia de este perro, posiblemente a través del cuidado humano (Janssens et al, 2018)
Evidencias genéticas
Un estudio del ADN mitocondrial de lobos y perros (C. Vilà, 1997) llegó a la conclusión de que los perros más antiguos podrían haberse originado hace 100 ka.Anna S. Druzhkova et al (2013) estudiaron el ADN-mt los restos de un cánido hallados en Altai, encontrando una relación más próxima con los perros domésticos que con los lobos.
Guo-dong Wang et al (2013) han secuenciado el genoma completo de varios lobos grises, perros chinos indígenas y perros de diversas razas. Según sus conclusiones, la división entre los lobos y los perros indígenas chinos se produjo hace 32 ka y los cuellos de botella posteriores fueron leves.
Adam H. Freedman et al (2013) analizaron el genoma de tres lobos grises, dos perros de linajes antiguos y un chacal. Los perros y los lobos divergieron por un proceso dinámico, con cuellos de botella de población en ambos linajes y flujo génico post-divergencia.
- En los perros, el cuello de botella implicó una reducción del tamaño de la población a la 17-49ava parte.
- En los lobos, el cuello de botella se produjo poco después de la divergencia con los perros.
- No se han conseguido indicios sobre el origen geográfico de la domesticación, pero en todo caso se pone en duda una procedencia del sudeste de Asia. Esto sugiere que el perro procede de una especie de lobo extinta.
- La posible fecha de la domesticación inicial del perro, queda en el rango de 16-11 ka, anterior al surgimiento de la agricultura.
- Los autores analizaron los datos genéticos de otras doce razas de perro y encontraron que el husky siberiano y el dingo, que no están relacionados con sociedades agrícolas, no tienen genes para la amilasa, que produce la digestión del almidón. En cambio estos genes sí están presentes en lobos.
Erik Axelsson et al (2013) han encontrado diez regiones genómicas en los perros
que tienen un papel clave en la digestión del almidón y las grasas
(genes AMY2B, MGAM y SGLT1) y que son diferentes a las correspondientes de
los lobos. Los perros coevolucionaron con el ser humano, la agricultura y la
correspondiente dieta.
O. Thalmann et al (2013) han estudiado el ADN-mt completo de 18 lobos prehistóricos y cánidos europeos y americanos y 77 perros, 49 lobos y 4 coyotes modernos, incluyendo las razas antiguas, como el Basenji, el Dingo, así como razas chinas. Para la calibración, tuvo en cuenta los fósiles de Goyet y Eliseevichi 1, considerándolos como perros, lo cual es al menos dudoso (Drake, Coquerelle y Colombeau, 2015).
Pontus Skoglund et al (2015) han secuenciado el genoma de un lobo de la península de Taimyr, en el norte de Siberia, datado en hace 35 ka.
O. Thalmann et al (2013) han estudiado el ADN-mt completo de 18 lobos prehistóricos y cánidos europeos y americanos y 77 perros, 49 lobos y 4 coyotes modernos, incluyendo las razas antiguas, como el Basenji, el Dingo, así como razas chinas. Para la calibración, tuvo en cuenta los fósiles de Goyet y Eliseevichi 1, considerándolos como perros, lo cual es al menos dudoso (Drake, Coquerelle y Colombeau, 2015).
- En los perros actuales se distinguen cuatro linajes genéticos con origen en Europa. El clado más numeroso se originó hace 18,8 ka. A este clado pertenece un ejemplar de una cueva de Suiza datado en 14,5 ka. Todos los perros actuales descienden de una hembra de hace 32,1 ka, mucho antes de la revolución agrícola.
- Según los autores, estos resultados implican que los perros domésticos son la culminación de un proceso que se inició con los cazadores-recolectores europeos y los cánidos con los que interactuaban: Los perros primitivos se habrían aprovechado de los cadáveres que dejaban atrás aquellos cazadores, y pudieron prestar ayuda a los humanos en la captura de presas o en la defensa frente a grandes depredadores.
- La mayoría de los perros actuales resultaron estar más estrechamente relacionados con los lobos antiguos que con los modernos. La población que dio origen a los perros modernos probablemente ya esté extinta.
- Es posible la existencia de otros procesos de domesticación que no hallan dejado descendencia.
Kelsey E. Witt et al (2014) han recopilado los estudios disponibles sobre el ADN-mt de perros nativos americanos y han secuenciado el correspondiente a 42 perros de yacimientos ubicados en Illinois, costa de la Columbia Británica y Colorado. De acuerdo con los resultados:
- La diversidad genética para algunas poblaciones es baja, consistente con las prácticas de cría deliberadas. La presencia de perros en las Américas puede datar de hace solo ca 10 ka, coincidente con la datación de la más antigua sepultura de perro hallada en América y muy posterior a la entrada del ser humano en el continente.
- Se han identificado cuatro nuevos haplotipos de ADNmt, así como múltiples haplotipos fundadores posibles. Algunos haplotipos se parecen mucho a los de los lobos, lo que sugiere una mezcla con lobos norteamericanos o tal vez una segunda domesticación de cánidos en las Américas.
- Las estimaciones iniciales de tamaño efectivo de la población sugieren al menos 1.000 hembras en vida del primer cánido conocido, y que el tamaño de la población aumentó gradualmente con el tiempo, antes de estabilizarse aproximadamente hace 1,2 ka.
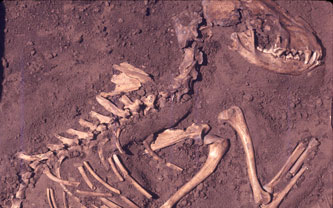 |
| Perro de Greene County III, Illinois, datado en 8,5 ka. (Del Baston, Center for American Archaeology) |
Este individuo pertenecía a una población que se separó del ancestro común de los lobos actuales y los perros muy poco antes de la aparición del linaje de los perros domésticos (trifurcación). Los autores han utilizado este genoma para recalibrar la tasa de mutación de perros y lobos, sustancialmente más lenta de lo previsto por la mayoría de los estudios anteriores.
Hay evidencia de introgresión del linaje del lobo de Taimyr en las actuales razas de perros del noreste de Siberia y Groenlandia.
Los perros domésticos se originaron Asia Suroriental hace 33 ka. Los perros de esta región tienen diversidad genética significativamente mayor y son el grupo más basal con relación a los lobos grises. Los perros indígenas chinos representan una forma intermedia entre los lobos y los perros de raza, y no han experimentado una selección artificial intensa. Mientras que la población de perros domésticos disminuyó drásticamente después de la división de la población, la de lobos experimentó un ligero crecimiento, posiblemente como consecuencia de las extinciones de la megafauna y con ello la reducción de competencia de otros depredadores.
 |
La población del lobo de Taimyr se separó del ancestro común de los lobos y los perros poco antes del origen del linaje de los perros domésticos. Hay introgresión genética del linaje del lobo de Taimyr en las razas actuales de perros del noreste de Siberia y Groenlandia. Pontus Skoglund et al (2015)
|
Guo-Dong Wang et al (2015) han secuenciado el genoma completo de 58 cánidos (12 lobos grises, 27 perros primitivos de Asia y África, y 19 de diferentes razas de todo el mundo).
De acuerdo con los resultados y tomando una tasa de mutación de 2,2 × 10-9 por sitio por año, la población ancestral de lobos comenzó a disminuir hace 2 Ma alcanzando un mínimo hace 400-300 ka, para aumentar luego alcanzando un máximo hace 200 ka.
De acuerdo con los resultados y tomando una tasa de mutación de 2,2 × 10-9 por sitio por año, la población ancestral de lobos comenzó a disminuir hace 2 Ma alcanzando un mínimo hace 400-300 ka, para aumentar luego alcanzando un máximo hace 200 ka.
Los perros domésticos se originaron Asia Suroriental hace 33 ka. Los perros de esta región tienen diversidad genética significativamente mayor y son el grupo más basal con relación a los lobos grises. Los perros indígenas chinos representan una forma intermedia entre los lobos y los perros de raza, y no han experimentado una selección artificial intensa. Mientras que la población de perros domésticos disminuyó drásticamente después de la división de la población, la de lobos experimentó un ligero crecimiento, posiblemente como consecuencia de las extinciones de la megafauna y con ello la reducción de competencia de otros depredadores.
Hace unos 15 ka, un subconjunto de los perros ancestrales comenzó la migración al Levante Mediterráneo, África y Europa. A este último continente llegó hace ca 10 ka. Uno de los linajes de Asia, viajó de vuelta hacia el este, creando en el Norte de China una serie de poblaciones endémicas mezcladas con los linajes asiáticos, antes de migrar al Nuevo Mundo. Una segunda oleada de migración hacia América se muestra en varias razas de perros de Sur y Centro América sin signos de mezcla con el linaje indígena del sur de China. Los perros de Australia y Nueva Guinea muestran fuertes señales de mezcla con lobos grises.
Laurent A. F. Frantz et al (2016) han secuenciado el genoma de un perro de la Edad del Bronce de Newgrange (Sí an Bhrú, Irlanda, hace ca 4,8 ka), y el ADNmt de 59 perros antiguos (entre hace 14-3 ka) y los han comparado con los de perros actuales de todo el mundo.
 |
| A. Evidencias arqueológicas de perros. B. Hipótesis de un doble origen de los perros |
Laurent A. F. Frantz et al (2016) han secuenciado el genoma de un perro de la Edad del Bronce de Newgrange (Sí an Bhrú, Irlanda, hace ca 4,8 ka), y el ADNmt de 59 perros antiguos (entre hace 14-3 ka) y los han comparado con los de perros actuales de todo el mundo.
De acuerdo con los resultados, los perros fueron domesticados a partir de dos poblaciones diferentes de lobos, del este y el oeste de Eurasia.
- La población occidental fue sustituida (al menos parcialmente) por perros del este de Eurasia.
 |
|
CTC: Perro procedente de Kirschbaumhöhle (Cherry Tree Cave), cerca de Forchheim, Alemania, datado en hace 2.900-2.632 ka cal
HXH: Perro procedente de Herxheim, Alemania, datado en hace 5.223-5.040 ka cal.
NGD: Perro irlandés estudiado por Laurent A. F. Frantz et al (2016).
|
Botigué et al (2017) han obtenido el genoma de un perro Neolítico temprano (HXH) y otro del Neolítico final (CTC), procedentes de Alemania y han incluido en el estudio una muestra asociada con una comunidad agrícola europea temprana.
- Ambos perros demuestran continuidad entre sí y comparten predominantemente ascendencia con perros europeos modernos, contradiciendo un reemplazo de la población en el Neolítico tardío.
- No han hallado ninguna evidencia genética para apoyar la hipótesis que propone un doble origen de la domesticación del perro.
- Mediante la calibración de la tasa de mutación, se estrecha el momento de la domesticación del perro a hace 40-20 ka.
Teorías sobre la domesticación
Las circunstancias exactas de domesticación han sido objeto de muchas especulaciones. Varias teorías se centran en el uso deliberado de lobos con fines prácticos: la caza, la vigilancia, o incluso como alimento. Sin embargo, es igualmente probable que la domesticación se produjera simplemente por accidente, por captura y cría de pequeños animales salvajes como mascotas.Los esqueletos de perros antiguos ponen de relieve el aplanamiento en el perro de las puntas de dorsales de las vértebras, lo que sugiere el transporte de pesadas mochilas a la espalda. Además, suelen faltar los molares cerca de la parte posterior de la mandíbula, lo que puede indicar que los animales llevaban algún tipo de brida para tirar de los carros.
Kent Flannery ha formulado la Hipótesis de la Revolución de Espectro Amplio (Broad Spectrum Revolution hypothesis, BSR) que afirma que la domesticación de animales y vegetales empezó en un periodo de carestía debida a condiciones climatológicas adversas, cuando los cazadores-recolectores tuvieron que ampliar su gama de fuentes de alimentos, incluyendo también los alimentos menos atractivos.
Friederike Range y Zsófia Virányi (2013) han comprobado que la habilidad de aprender no solo de sus congéneres sin también de los humanos está presente en el lobo.
De acuerdo con la Hipótesis de la reactividad emocional (Hare y Tomasello, 2005) la selección para un temperamento más doméstico y la reducción del miedo y la agresión, explica el mayor éxito de los perros en las interacciones de cooperación y de comunicación con los seres humanos en comparación con los lobos. Según Hare et al (2012), durante la domesticación los perros se volvieron menos agresivo y más tolerantes que los lobos no sólo hacia los seres humanos, sino también hacia sus congéneres (Virányi y Range, 2014).
 |
| Hipótesis de la Cooperación Canina. Range y Virányi, 2015. |
La Hipótesis de la Cooperación Canina (Range y Virányi, 2013, 2014; Virányi y Range, 2014; Range y Virányi, 2015) sugiere por el contrario que los lobos se caracterizan por su alta atención social y tolerancia y que son muy cooperativos. Estas características de lobos probablemente constituían una buena base para la evolución de la cooperación entre perros y humanos.
Los cerdos fueron domesticados de forma independiente en el Este de Asia (Cucchi et al, 2011) y Anatolia (Ottoni et al, 2013).
- A diferencia de muchas hipótesis de domesticación, esta hipótesis sugiere que los perros no fueron seleccionados por un aumento general de su atención social y tolerancia. Cuando se les proporciona socialización humana intensiva, los lobos aceptan los seres humanos como interlocutores sociales y pueden utilizar la información de las personas como lo hacen los perros.
- La Hipótesis de la Cooperación Canina es compatible con otras teorías evolutivas que enfatizan el papel que jugaron en la evolución de los perros las similitudes entre los lobos y los humanos en la sociabilidad y la cooperatividad (Schaller y Lowther, 1969; Clutton-Brock, 1984; Schleidt, 1998) y con las hipótesis de domesticación que abordan específicamente la conducta de perros dirigida a humanos. Por ejemplo, se ha sugerido que se seleccionaron perros por el reducido recelo y fácil socialización con los seres humanos (Scott y Fuller, 1965; Klinghammer y Goodmann, 1987); esto sin duda les ayudó a ampliar sus habilidades sociales heredadas de los lobos.
 |
| Cladograma de perros y lobos. Anna S. Druzhkova el al (2013). |
BÓVIDOS Y SUIDOS
Se cree que varias especies de bóvidos fueron domesticadas en diferentes lugares, como Asia suroccidental y el norte de China, hace más de 10 ka (Hucai Zhang et al, 2013).Los cerdos fueron domesticados de forma independiente en el Este de Asia (Cucchi et al, 2011) y Anatolia (Ottoni et al, 2013).
 |
| Geografía de la domesticación de los bóvidos. Hucai Zhang. |
Marom y Bar-Oz (2013) han publicado un estudio de la interacción humana con los bóvidos y los suidos (Bos taurus y Sus scrofa) en el Valle del Jordán. Seguramente la domesticación de estos animales se realizó en múltiples ocasiones y lugares. En el norte del Valle del Jordán, la incorporación de bóvidos y suidos sucedió hace 8 ka.
La sobrecaza y disminución de las poblaciones salvajes, precedió a la domesticación. La selección natural actuó sobre estas especies con los siguientes efectos:
- Disminución de tamaño.
- Aumento de la tasa de rotación, con una edad más joven para la procreación.
- Neotenia.
Además, la eliminación de los individuos más grandes y agresivos, permitió a los humanos tomar su papel como líderes en la estructura jerárquica.
Jared E. Decker et al (2013) han estudiado el genoma de 1.543 bóvidos domésticos de 134 razas de todo el mundo. Estas razas surgieron de tres subespecies domesticadas: Bos javanicus, Bos taurus indicus (valle del Indo) y Bos taurus taurus (Creciente Fértil).
Jared E. Decker et al (2013) han estudiado el genoma de 1.543 bóvidos domésticos de 134 razas de todo el mundo. Estas razas surgieron de tres subespecies domesticadas: Bos javanicus, Bos taurus indicus (valle del Indo) y Bos taurus taurus (Creciente Fértil).
- Los bóvidos africanos surgen del cruce del uro salvaje africano con el Bos t. taurus domesticado en el Creciente Fértil.
- Los bóvidos euroasiáticos y americanos proceden del cruce de Bos t. taurus, sin joroba, y de Bos t. indicus, con joroba, para producir híbridos adaptados a los sistemas de producción y al medio ambiente locales (Upadhyay et al, 2017).
- La historia del ganado de Asia implica la domesticación y la mezcla de varias especies. Además de ganado domesticado de uro (Bos primigenius), también contribuyeron al acervo genético el búfalo de agua domesticado (Bubalus bubalis), yak (Bos grunniens), gaur (Bos gaurus) y banteng (Bos javanicus).
- Las razas de Anatolia provienen de una mezcla de ganado europeo, africano y asiático y no representa a las poblaciones originalmente domesticadas en esta región.
- La raza Zavot muestra una historia diferente, con una gran parte de ancestros similares a Holstein.
- El ganado vacuno del norte y noroeste de Asia desciende principalmente de Bos t. taurus, pero las razas Hanwoo y Mongol también ascienden de Bos t. indicus.
- El ganado de Pakistán, India, sur de China e Indonesia asciende predominantemente de Bos t. indicus.
- El ganado entre estas dos regiones geográficas es híbrido de Bos t. taurus × Bos t. indicus.
- Las razas Pesisir, Aceh, Hainan, Luxi, Brebes y Madura muestran una ascendencia significativa de Bos javanicus.
- En la formación de las razas europeas, se detecta la influencia pronunciada del ganado Shorthorn y la mezcla con poblaciones silvestres (Upadhyay et al, 2017). El ganado fue penetrado de sureste a noroeste.
- Según Jörg Schibler, Julia Elsner y Angela Schlumbaum (2014) los ganaderos europeos cruzaron uros hembra (Bos primigenius) con toros domésticos.
- Park et al (2015) y Upadhyay et al (2017), para el ganado británico e irlandés. El ganado importado se mezcló con uros locales.
- El ganado ibérico e itálico posee introgresión de Bos t. taurinus africano.
- Tres razas italianas conservan la presencia de Bos t. indicus, lo cual es consistente con dos migraciones separadas, una de Asia Occidental, donde se había producido introgresión de Bos t. indicus y otra de África Occidental, pasando por España, sin esta ingrogresión. Este segundo grupo parece haber irradiado desde España por el Mediterráneo europeo.
- Los bovinos criollos estadounidenses son de procedencia ibérica y no africana. Tras la llegada a América, se produjo mezcla con ganado importado desde la India.
 |
| Una mutación permitió la asimilación de la lactasa y con ello el desarrollo de una economía basada en los productos lácteos. Andrew Curry, 2013. |
 |
| Camino a la domesticación del ganado bovino y porcino en el Norte del Valle del Jordán. |
CABALLOS
Los caballos fueron domesticados por primera vez en un único
lugar de las estepas que actualmente ocupan los países de Ucrania, Rusia y
Kazajstán, hace unos 5,5 ka.
Dogs: Scientist’s Best Friend (Science Online Extra). 2015.
Según Ludovic Orlando et al (2013), las poblaciones de caballos domésticos y el caballo de Przewalski divergieron hace 72-38 ka y han permanecido aisladas.
Los caballos de Przewalski representan los últimos supervivientes de las poblaciones salvajes. Conservan una alta variación genética.
A lo largo de la domesticación, fueron seleccionados positivamente genes implicados en diversos aspectos de la locomoción, la fisiología, y la cognición. El costo genético de este proceso está representado por un exceso de mutaciones deletéreas en los caballos modernos (Mikkel Schubert et al, 2014)
A lo largo de la domesticación, fueron seleccionados positivamente genes implicados en diversos aspectos de la locomoción, la fisiología, y la cognición. El costo genético de este proceso está representado por un exceso de mutaciones deletéreas en los caballos modernos (Mikkel Schubert et al, 2014)
 |
| Caballos Przewalski en Khomyntal, Mongolia occidental, uno de los tres sitios de reintroducción. Los caballos fueron importados en 2004 de Le Villaret, Francia. Imagen de Claudia Feh, Association pour le cheval de Przewalski. |
Mecanismos genéticos de la domesticación
La domesticación de animales salvajes induce un conjunto de características fenotípicas conocidas
como síndrome de domesticación. La hipótesis del déficit de células de la cresta neural propone que se genera por un programa de desarrollo ligeramente interrumpido.
A partir de una investigación sobre el proceso de domesticación de las lubinas, Anastasiadi y Piferrer (2019) han argumentado que las etapas iniciales de la domesticación incluyen alteraciones dinámicas en la metilación del ADN de los genes del desarrollo que afectan la cresta neural (NCC), epimutaciones que podrían fijarse como variantes genéticas.
Enlaces:
Dog genetics spur scientific spat. Ewen Callaway. 2013.
Dogs: Scientist’s Best Friend (Science Online Extra). 2015.