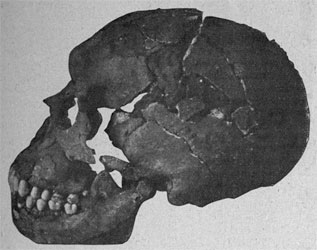
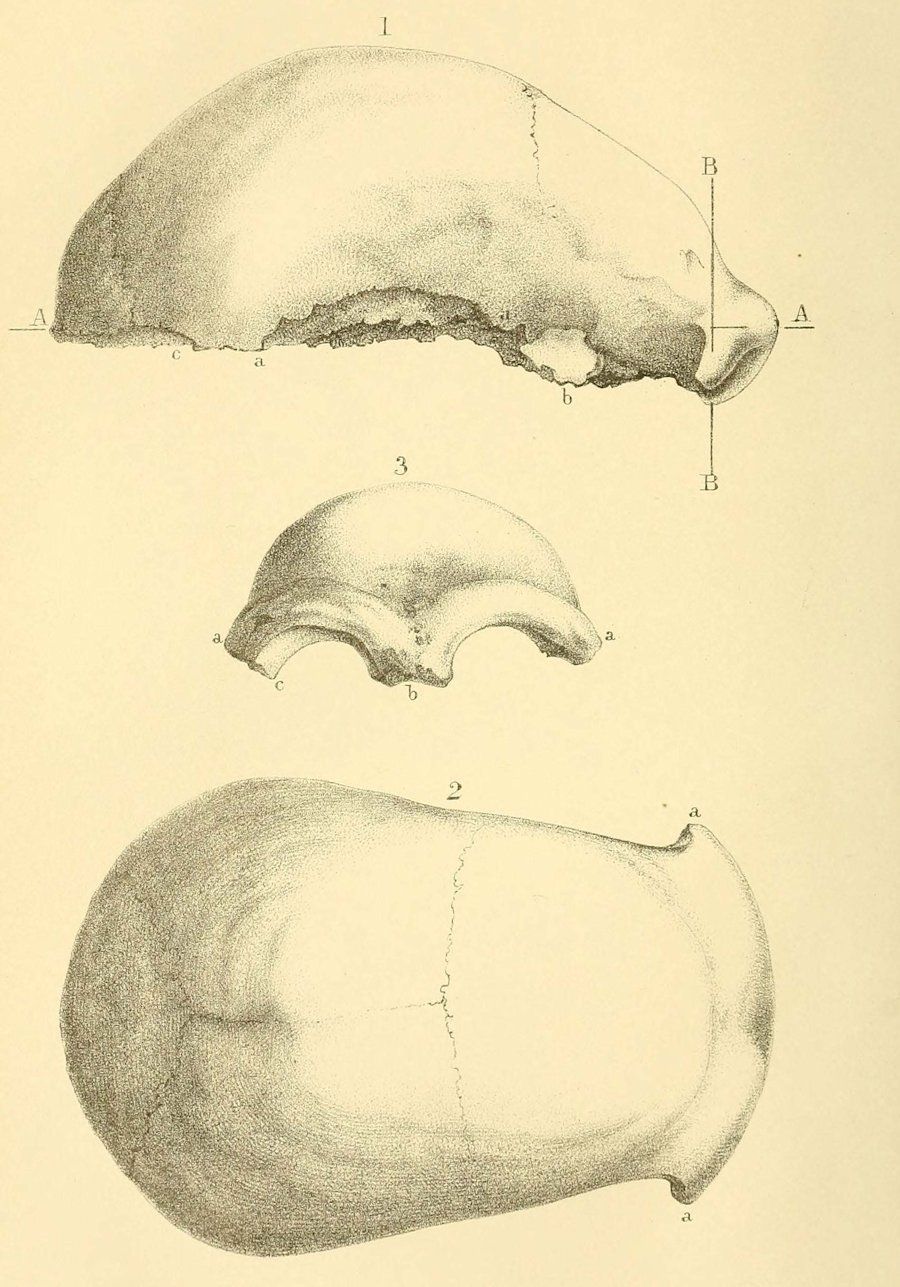
Homo floresiensis, Brown et al, 2004
 |
| LB 1. Holotipo de Homo floresiensis |
- Gerrit D. van den Bergh et al (2016) han comunicado el hallazgo de 41 artefactos líticos in situ asociados con restos fósiles de megafauna (Bubalus sp., Stegodon y Celebochoerus) en la trencha 4 (T4) y otros cuatro de pequeño tamaño en la sub-unidad E2, más antigua, en Talepu, Cuenca Walanae, noreste de Maros, isla de Sulawesi. Los artefactos no han podido ser clasificados en ninguna industria en particular. La edad mínima es de hace 118 ka mientras que los artefactos de E2 pueden llegar hasta hace 194 ka.
 |
| Posibles rutas hasta Flores de los antepasados del Homo floresiensis |
- LB1: Gran parte del esqueleto de una hembra adulta. Incluye el pie izquierdo relativamente completo y partes del derecho. Fue datado en 18.000 años, pero un nuevo estudio estratigráfico ha proporcionado una edad de hace 100-60 ka, mientras que para los artefactos la datación es de hace 190-50 ka (Sutikna et al, 2016; Sutikna et al, 2018)
- LB2: Un premolar inferior de otro individuo.
Este descubrimiento planteó una paradoja: Con un cerebro de tamaño parecido al de un chimpancé, la nueva especie mostraba indicios de inteligencia avanzada como por ejemplo la caza con armas de piedra, despedazar cadáveres o controlar el fuego.
- La hipótesis del sesgo de cautividad relaciona la frecuencia de uso y variedad de las herramientas con el tamaño y la interconexión de las poblaciones y no con la capacidad cognitiva (Michael Haslam, 2013).
 |
| LB1. Homo floresiensis. Reconstrucción de la cara. Susan Hayes et al (2013). |
- LB3: un cúbito.
- LB4: radio y tibia de un niño.
- LB5: vértebra y metacarpo de un adulto.
- LB6: algunos huesos de la mano, omóplato y mandíbula. Esta mandíbula es muy similar a la de LB1.
- LB7: hueso del pulgar.
- LB8; tibia.
- LB9: fémur.
 |
| Hallazgos de Homo floresiensis |
- LB11 incluye fragmentos de pelvis y un metatarso fragmentario.
- LB12, falange de la mano, distal.
- LB13 es un fragmento de la rótula.
- LB14 es un fragmento de un acetábulo.
- LB15/1, cuarto premolar inferior.
- LB15/2, incisivo superior.
- LB16, cuboide del pie derecho.
- LB20, LB21 y LB22, huesos del carpo carentes de las características derivadas presentes en neandertales y sapiens.
La secuencia estratigráfica de la cueva está bien definida, pero comprende sedimentos interestratificados de diversas litologías y orígenes, incluyendo tefras volcánicas, colluvium de grano fino, gravas calizas autógenas gruesas, espeleotemas y sedimentos antropogénicos, como restos de combustión. La heterogeneidad sedimentológica y química sugiere que los procesos de formación de sitio y diagénesis variaron notablemente a través del tiempo, tanto lateral como verticalmente (Morley et al, 2016).
a |
| Estratigrafía de Liang Bua. A la izquierda, Sutikna et al, 2016. A la derecha, Morley et al, 2016. |
En 2014 se recuperaron en Mata Menge, Cuenca del So'a, en el centro de la Isla de Flores, restos pertenecientes al menos a tres individuos.
- Un fragmento de mandíbula, de un adulto.
- 6 dientes aislados. Dos caninos corresponden a dos diferentes individuos infantiles.
Los ejemplares son similares en dimensiones (incluso más pequeños) y características morfológicas a los hallazgos de Liang Bua, salvo en el caso de un primer molar mandibular, que retiene una condición más primitiva (van den Berg et al, 2016). Los fósiles fueron hallados entre sedimentos fluviales de piedra arenisca, depositados hace ca 700 ka, de acuerdo con los resultados de los diferentes métodos utilizados. Los datos paleoambientales indican un clima relativamente seco y un hábitat de pradera abierta con humedales. El conjunto lítico asociado responde a una tecnología simple notablemente similar a la de Liang Bua (Brumm et al, 2016).
Según María Martinón-Torres et al (2017), las evidencias no son suficientes para afirmar una relación ancestro descenciente entre los individuos de Manta Menge y los de Liang Bua. Mata Menge podría representar uno de los varios episodios de ocupación de Flores e islas cercanas, confirmando la naturaleza compleja de los intercambios intermitentes que pueden haber ocurrido entre el sudeste asiático continental e insular durante todo el Pleistoceno.
En la cueva de Liang Bawah, a la que se accede desde Liang Bua por un pasillo descendente. se hallaron 17 artefactos de piedra y materiales volcánicos (Gagan et al, 2015).Según María Martinón-Torres et al (2017), las evidencias no son suficientes para afirmar una relación ancestro descenciente entre los individuos de Manta Menge y los de Liang Bua. Mata Menge podría representar uno de los varios episodios de ocupación de Flores e islas cercanas, confirmando la naturaleza compleja de los intercambios intermitentes que pueden haber ocurrido entre el sudeste asiático continental e insular durante todo el Pleistoceno.
 |
| Localización de las cuevas Liang Bua y Liang Bawah, en la Isla de Flores (Gagan et al, 2015). |
Floresiensis se define por:
- Estatura sobre 1,06 m. 32,5 kg.
- Muy pequeña capacidad craneal (426 cc; Daisuke Kubo et al, 2013). Bóveda craneana más gruesa que en Australopitecus y similar a H. erectus y H. sapiens. La bóveda craneana es baja y angosta en comparación con la del hombre actual y aun en comparación con la de erectus. La flexión basicraneal que es un rasgo determinante en la hominización, es de 130°, intermedia entre erectus (141°) y el hombre actual (127°). Su estatura total y su volumen craneano ofrecen valores que están por fuera del rango de variación de los homínidos modernos. Coincidentemente, el grado de encefalización –expresado como cociente entre el volumen endocraneano y la longitud femoral (gran indicadora, esta última, de la talla total)– arroja para floresiensis un valor (5,9) menor al de erectus (6,9) y aún menor al de afarensis (6,1), mientras que en los homínidos actuales dicho valor gira en torno a 7,5. Para Dean Falk, los rasgos cerebrales preservados en su cráneo sugieren que el homínido de Flores podría ser capaz de realizar tareas cognitivas avanzadas.
- El cerebro del Homo floriensis posee lóbulos temporales bastante amplios, regiones del cerebro asociadas con la comprensión del habla y el oído. Aún más sorprendente es el hecho de que posee lóbulos frontales del cerebro que tienen circunvoluciones con muchos pliegues, áreas del cerebro directamente debajo de la frente que están implicados en un reconocimiento superior. La región más circunvolucionada es la parte que se proyecta más allá del lóbulo frontal, llamada el polo frontal. Falk identifica esta región como área 10 de Brodmann, la cual está expandida en humanos modernos y está relacionada con la toma de iniciativas e ideación de planes de acciones futuras que son componentes clave de la cognición elevada. Esta área alargada sugiere que Homo floresiensis pudo confeccionar las herramientas de piedra que se encontraron en las proximidades, las cuales son más típicas de humanos prehistóricos modernos que de antiguos homínidos como erectus.
- Se parece al subadulto de Dmanisi D2700. El grosor de los huesos craneales recuerda también a H. erectus.
- En la comparación de 67 rasgos craneofaciales, Kaifu et al (2011) interpretaron que el patrón de floresiensis es consistente con una ascendencia de los erectus tempranos de Indonesia. Sin embargo, identificaron al menos 20 rasgos craneales de LB1 que también se encuentran en habilis y algunas poblaciones de erectus, incluyendo un débil o nulo desarrollo de las quillas coronal y sagital, un escama temporal grande con una sutura parietomastoidea corta, la presencia de un proceso postglenoideo, una fosa mandibular poco profunda y un surco supraorbital que no se distingue con claridad.
- Arcos superciliares de tamaño moderado.
- Mandíbulas salientes. La sínfisis mandibular no se resuelve en un verdadero mentón como en sapiens, sino que presenta inclinación en sentido póstero-inferior del eje sinfisario, como en los australopitecinos o el habilis. La fuerte inclinación de la sínfisis mandibular es compartida con algunos ejemplares de la población de Dmanisi, erectus y australopitecinos gráciles, pero no con los primeros Homo ni con los erectus de Indonesia. Protuberancias o contrafuertes mandibulares, comunes en los homínidos tempranos pero no en el HAM. Hueco entre el extremo de la fila de dientes y la sección posterior de la mandíbula. La forma mandibular general ha sido descrita como similar a la de los primeros erectus africanos y a la población de Dmanisi, aunque con algunas características más primitivas. La prominencia alveolar lingual bien desarrollada es compartida únicamente con los erectus africanos.
- Sin las especializaciones masticatorias ya presentes en Australopithecus y Paranthropus. Muelas relativamente grandes, aunque las coronas molares mandibulares son más pequeñas que en de los primeros Homo y australopitecinos. Raíces dentales diferentes al HAM.
- Según David J. Daegling et al (2014) la mandíbula de floresiensis es relativamente fuerte (comparable a la de los australopitecinos) en términos de rigidez en la torsión y la flexión transversal, pero es relativamente débil bajo flexión parasagital. Las cargas masticatorias se redujeron en relación con los australopitecinos, pero se mantuvieron relativamente elevadas con respecto a los HAM.
- La escala M1> M2> M3 es compartida con erectus y sapiens peron no con los primeros Homo.
- Yousuke Kaifu et al (2015) han analizado los restos dentales de Homo floresiensis.
- Conjunto C/P3 y otras características primitivas, similares a Homo erectus.
- C1: hombro distal bajo.
- P3: cresta transversal. Esmalte bucal hinchado (cíngulo). La orientación de la cresta transversal distolingual en el P3 inferior es una característica derivada compartida con Dmanisi y los erectus africanos. La corona asimétrica y alargada mesiodistalmente del P3 inferior, documentada en LB1 y LB6 es común en los australopitecinos y en Homo temprano y, aunque es rara en erectus, se observa en Dmanisi. La raíz bifurcada es común en los australopitecinos y se observa también en algunos erectus tempranos.
- P4: Cresta transversal. Cara lingual de la corona, bien desarrollada. LB1 presenta un P4 inferior con la raíz dividida. Este tipo de raíz (raíz de Tomes), con la cámara pulpar simple en forma de cinta arqueada 180º o más (C-shaped) aparece por vez primera en habilis coincidiendo con la reducción en tamaño de los dientes, pero mientras que para los P3 inferiores se han documentado en los primeros Homo, en los P4 inferiores son poco comunes en el registro fósil, pero están presentes en los primeros Homo y en los erectus de África e Indonesia.
- M1: cresta media del trigónido.
- Tamaños equivalentes de M1 y M2.
- Morfología molar avanzada, incluso en comparación con los HAM.
- Apomorfias de H. floresiensis:
- Gran tamaño relativo y morfología oclusal única del P3.
- Primeros molares extremadamente pequeños. No está clara la razón por la que H. floresiensis experimentó una evolución tan singular.
- Esta combinación de rasgos es desconocida hasta el momento en otras especies de Homo. La dentición de H. floresiensis no es una versión a escala reducida de Homo anteriores.
- Prognatismo facial. Frente huidiza.
- Tiene arcos supraorbitarios marcados, aunque no llega a formar el clásico torus.
- Meatus auditivo externo pequeño y redondeado, algo alargado verticalmente, con margen timpánico muy fino, que recuerda a los australopitecinos.
- Bajos ángulos glenoideo y axilo espinal del omóplato. Recuerda a Turkana boy (ergaster) y australopitecinos.
- LB1 / LB6 comparten baja torsión humeral y bajo ratio claviculohumeral con erectus y Hominini anteriores.
- Clavícula corta.
- Muñeca primitiva, que recuerda la de los australopitecinos.
- Trapezoide acunado.
- El lado cubitar de la articulación del segundo metacarpo, orientado sagitalmente
- La articulación del escafoide, algo triangular
- Falta el área de expansión palmar característica del último Homo y del Pleistoceno Medio, que permite la pinza de precisión.
- Escafoide y capitate plesiomórficos. Capitate con un cuello entallado y una faceta del escafoides que se extiende sobre el cuello en forma de J.
- Piernas cortas, como las de los australopitecinos.
- En KNM-WT 15000 y LB6, la fosa glenoidea se orienta lateralmente en lugar de cranealmente, lo que constituye una morfología derivada desconocida en los primeros Homo. En Dmanisi en cambio, se orienta más cranealmente.
- La poca profundidad dorsal del surco troclear de LB1, es una condición derivada en la dirección del sapiens que aparece tanto en los Homo tempranos como en los erectus.
- Las alas abiertas hacia fuera en la faceta del maleolo externo parecen conservarse en la mayor parte de los fósiles anteriores al sapiens.
- Pie excepcionalmente largo en relación al fémur y a la tibia. Junto con otras características, los rasgos del pie indican una forma de desplazamiento biomecánicamente distinta a la del sapiens (W. L. Jungers et al, 2009).
- La prominente tuberosidad navicular de LB1 es más primitiva que la observada en OH8 y parece indicar que carecía de arco longitudinal medial, aunque también Jinniushan y los neandertales muestran una tuberosidad navicular prominente, por lo que la interpretación es controvertida.
- El bajo grado de torsión de la cabeza del astrágalo en LB1 está más cerca de los monos africanos que de los humanos, aunque se han reportado valores similares para OH8.
- El hállux es bastante corto en relación a los otros dedos, lo que constituye una característica primitiva, similar a la que aparece en Dmanisi. En LB1, Dmanisi y los Hominini de Illeret y está aducido al resto del dedos en contraste con los Hominini basales y Burtele o el grado intermedio documentado para los australopitecinos. Sin embargo, las huellas de Laetoli y OH8 también muestran un hállux aducido.
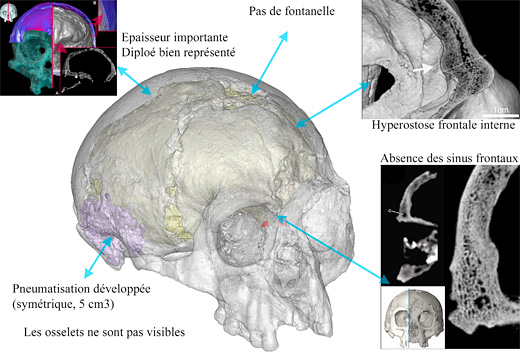 |
| Características craneales de LB1. © Antoine Balzeau-CNRS/MNHN. |
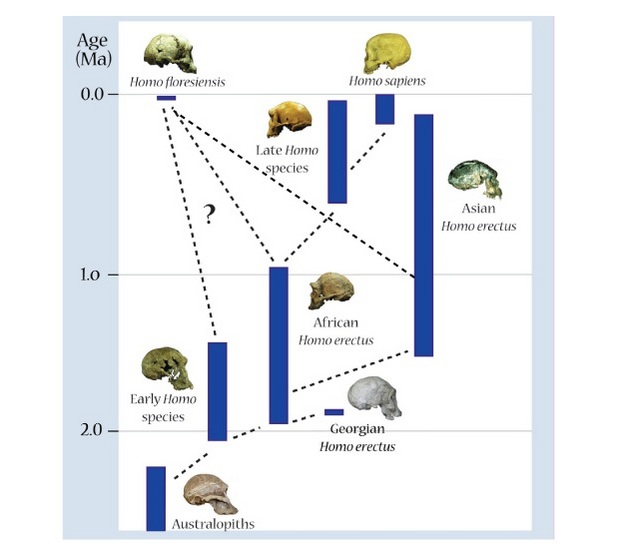 |
| Posibles escenarios evolutivos para Homo floresiensis |
Teorías sobre el origen del Homo floresiensis
Según el análisis cladístico efectuado por D. Argue et al (2012), los resultados producen dos árboles filogenéticos igualmente parsimoniosos. El primero sugiere que H. floresiensis es un homínido temprano que surgió después de Homo rudolfensis, pero antes de que H. habilis. El segundo árbol indica que H. floresiensis surgió después de Homo habilis.
- Homo floresiensis no es humano.
- Alfred Czarnetzki (2014) propone que floresiensis es una subespecie de Pongo abelii.
- Los restos hallados corresponden a casos patológicos. Estas propuestas no explican como se pudo mantener la patología durante tantos miles de años y la ausencia de especímenes sin la referida patología.
- Oxnard et al (2013) han criticado los estudios y conclusiones de Brown et al, quienes consideran que floresiensis fue un descendiente de erectus, en una línea evolutiva afectada por el enanismo insular
- Para Teuku Jacob y Robert Martin, LB1 se trataba de un sapiens que padeció microcefalia. Oxnard et al, 2010, Brown 2012 los consideran casos de cretinismo.
- Opinan lo mismo Obendorf et al (2008) en base al tamaño de la silla turca y Vannucci el al (2011) en base a un análisis de tomografía computerizada.
- Para Falk et al. 2009, (ver Hawks 2007), estamos ante un caso de síndrome de Laron.
- Robert B. Eckhardt et al (2014), Robert B. Eckhardt et al (2015), consideran que las principales características (volumen cerebral y estatura) de LB1 están subestimadas ya que se ha pasado por alto la asimetría y la desproporción del ejemplar, que son signos de desarrollo anormal. El ADN extraído de los huesos de LB1 es demasiado fragmentario para permitir un diagnóstico, pero los fragmentos recuperados hasta el momento son de tipo humano moderno. Según los autores, incluso en ausencia de un diagnóstico sindrómico en particular, las características que definen a LB1 no cumplen con los criterios formales para un espécimen tipo de una nueva especie.
- Para Maciej Henneberg et al (2014), la asimetría facial, el pequeño volumen endocraneal, la braquicefalia, los fémures desproporcionadamente cortos, los pies planos, y otras numerosas características de LB1 son altamente diagnósticas de síndrome de Down, uno de los trastornos del desarrollo que aparecen más frecuentemente en los seres humanos y también se documenta en los hominoideos como los chimpancés y los orangutanes. Consideran que el mentón, cuya presencia es un importante caracterísitica definitoria del HAM, está a menudo ausente entre los australo-melanesios.
- Michael Carrington Westaway et al (2015) advierten que los estudios sobre la ausencia de mentón de los autralo-melanesios no han sido revisados o han sido criticados severamente y consideran, de acuerdo con Brown y Tomoko, que la ausencia de barbilla en LB1 y LB6 imposibilita su atribución a H. sapiens. Otras características de la barbilla confirman esta imposibilidad.
- Para Baab et al (2016) la presencia en LB1 de características clínicas del síndrome de Down, es mínima.
- La datación de Sutikna et al (2016) de hace 100-60 ka, y el hallazgo de los fósiles de Mata Menge, datados en 700 ka, son poco conciliables con esta hipótesis.
- Según Antoine Balzeau y Philippe Charlier (2016) el cráneo LB1 no muestra ningún rasgo apomórfico de sapiens y su espesor no es compatible con una atribución a esta especie. Tampoco hay evidencia de patología sistémica.
- H. floresiensis experimentó una dramática reducción de tamaño del cerebro y del cuerpo a partir de la condición de grado erectus en un entorno insular aislado (enanismo insular). Según Kubo et al (2013), si H. floresiensis descendiese del erectus de Java (860 cc), al menos un 50% de la reducción de tamaño del encéfalo vendría explicada por la reducción del tamaño del cuerpo. Si descendiese de Homo habilis (600 cc), la reducción alométrica explicaría el pequeño tamaño del encéfalo en un porcentaje mucho mayor.
- Una primera hipótesis explicativa recurrió al enanismo insular (dwarfism), fenómeno evolutivo observado en las islas en muchas ocasiones consistente en una gran reducción de tamaño, como forma de sobrevivir en un entorno pobre. Para Peter Brown y colegas, floresiensis fue un descendiente de erectus, en una línea evolutiva afectada por el enanismo insular.
- Hay detalles de la antomía de LB1 que recuerdan al erectus: El neurocráneo, la frente plana e inclinada, el espesor craneal de los huesos, la cara corta y plana, el toro occipital, la fisura del mastoidos y el hombro, pero tanto el pequeño tamaño del cuerpo y el tamaño del cerebro se encuentran fuera del rango esperado para la especie (Brown et al, 2004; Falk et al, 2005; Baab y McNulty, 2009; Baab, McNulty y Harvati, 2013). Igualmente, la dentición es similar a la de erectus (Kaifu et al, 2015). Zeitoun et al (2016) llegaron a la conclusión de que LB1 se encuentra dentro del rango de variación de los primeros erectus de Java. Para Dembo et al (2015), H. floresiensis está incluido en el mismo clado que H. rudolfensis, H. erectus y las especies posteriores de Homo (pero no H. habilis).
- Según esta teoría, los ancestros de los Homo floresiensis de alguna manera navegaron hasta Flores, donde se produjo una reducción en el tamaño del cuerpo (Meijer et al, 2010). Se ha especulado con una migración no documentada hasta el momento, de erectus tempranos, relacionados con Dmanisi, de tamaño relativamente pequeño, al sudeste de Asia, de la que provendría floresiensis.
- Las características primitivas, las morfologías más similares a los australopitecinos que a Homo, han de corresponder a convergencias evolutivas. Tal vez la condición primitiva de la rama y la sínfisis mandibular se deba a un cambio hacia una masticación más poderosa. La morfología de la tuberosidad navicular, la ausencia de arco longitudinal y el pequeño tamaño del cerebro son otras reversiones tanto con respecto a erectus como a Homo temprano.
- Aumentó la robustez relativa de las extremidades superiores, lo que permite especular con la presencia de comportamientos más arbóreos, si bien las características del pie no apoyan esta hipótesis (Baab, 2016).
- La edad de los fósiles de Mata Menge (hace 700 ka) permite especular con un proceso de enanismo rápido (van den Bergh et al, 2016)
- H. floresiensis deriva directamente de una forma más primitiva de encéfalo pequeño como el Homo habilis (aprox. 600 cc) o incluso Australopithecus (aprox. 400 cc).
- No existe ninguna evidencia fósil en Asia de estas especies.
- Las evidencias craneales, de la mandíbula, los dientes y el esqueleto postcraneal apoyan esta hipótesis y los análisis filogenéticos son consistentes con un taxón hermano de habilis o con un taxón basal anterior (Argue et al, 2009; Argue et al, 2017). El tamaño y la morfología de los dientes y la mandíbula comparten más semejanzas con Australopithecus y los primeros Homo que con los erectus (Brown y Maeda, 2009). En particular, las piernas muy cortas (con respecto tanto a los brazos como a los pies) son un patrón común en los monos y los australopitecinos pero no en erectus. LB1 también era desproporcionadamente pesado para su altura, como Lucy (Australopithecus afarensis) (Jungers y Baab, 2009). Los huesos del carpo (muñeca) se parecen más a los de los chimpancés a los de los humanos modernos (Tocheri et al, 2007). La morfología del pie, también conserva varias características muy primitivas: una larga parte delantera del pie con los dedos curvados y la falta de un arco longitudinal medial (Jungers et al, 2009).
- H . floresiensis no sigue la tendencia alométrica del Homo del Pleistoceno Medio, los neandertales y el HAM (Juan Antonio Pérez-Claros, Juan Manuel Jiménez-Arenas y Paul Palmqvist, 2015)
- Si H. floresiensis derivó del HAM en condiciones insulares, tuvo que ser a través de transposiciones laterales. Esto significa que debe ser pedomórfico en tamaño y peramórfico en forma con respecto al HAM.
- Sin embargo, es más fácil conectar LB1 con los habilinos por escalamiento ontogénico, lo que significa que LB1 sería pedomórfico tanto en tamaño como en forma. Esta es una explicación más parsimoniosa.
- Si floresiensis es descendiente de habilis, muchas características del cráneo y algunos rasgos dentales tuvieron que evolucionar en paralelo en floresiensis y erectus (homoplasias; Baab, 2016). Algunos de estos rasgos pudieron evolucionar en forma correlacionada, lo que deja abierta la posibilidad de que los rasgos compartidos surgieran gracias a la selección de uno o unos pocos en presencia de presiones similares. Si OH 62 midió unos 118 cm, la estatura de LB1 de unos 106 cm requirió solo una pequeña reducción.
- Esta ascendencia supondría una migración Out of Africa no documentada hasta el momento.
- Es el resultado de un grupo enano de H. erectus con algún grado de introgresión de H. sapiens, aunque no se puede descartar el aporte de una especie diferente. Esto explicaría las anomalías dentales, craneales y postcraneales que se han utilizado durante mucho tiempo para cuestionar su estado taxonómico (María Martinón-Torres et al, 2017).
- El P3 del floresiensis es inusual en términos de forma y tamaño absoluto y relativo con respecto a otros elementos de la dentición. Además, la forma de los molares inferiores también es atípica (Kaifu et al, 2015). Más allá de la expresión de una cresta de midtrigónida, el patrón de crestas adicionales en la unión esmalte-dentina no se ajusta a ninguno de los patrones descritos hasta ahora en humanos extintos y existentes. Las patologías dentales que muestran los dientes, pueden ser consecuencia de una hibridación.
- Kaifu et al (2015), sugirieron que las características molares avanzadas de Liang Bua podrían ser el resultado de restricciones de espacio en las mandíbulas de H. floresiensis. Alternativamente, podrían ser el resultado del flujo de genes de H. sapiens. Los rasgos molares derivados de H. floresiensis también se han identificado en una muestra de Sangiran reportada por Zanolli (2013) y podría ser un indicador de afinidad biológica.
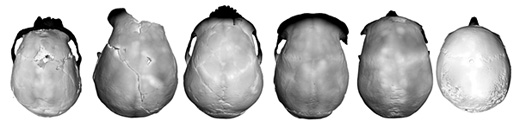 |
| De izquierda a derecha, LB1 (Homo floresiensis), Sangiran 2 (Homo erectus), La Ferrassie 1 (Homo neanderthalensis), Broken Hill (Homo rhodesiensis), Cro-Magnon 1 y un Homo sapiens con microcefalia actual. © Antoine Balzeau-CNRS/MNHN |
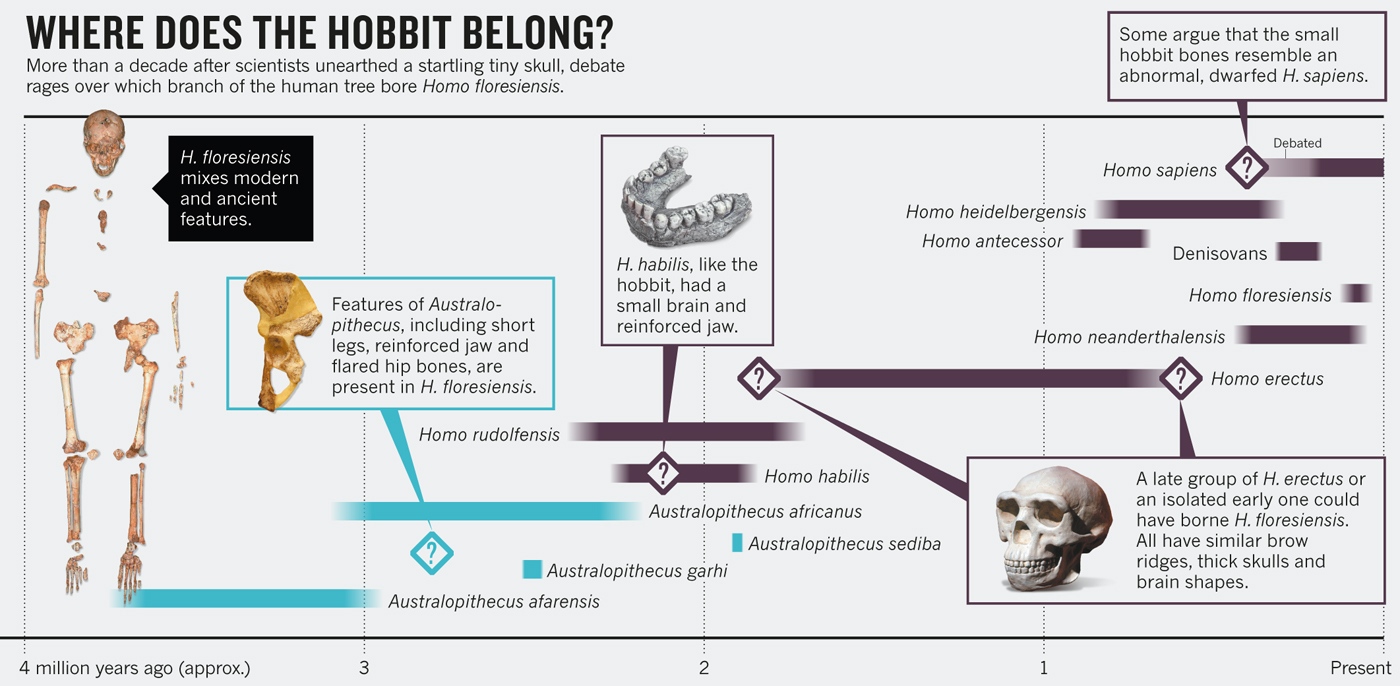 |
| Teorías sobre el origen de Homo floresiensis |
El hábitat del floresiensis
Hanneke J. M. Meijer et al (2013) han publicado un estudio de los restos de aves recuperados en el sector XI de la cueva de Liang Bua, donde se halló en 2003 el Homo floresiensis. El conjunto está formado por 244 muestras pertenecientes al menos a 26 taxones no paseriformes. Las especies representadas sugieren que durante los últimos 95 ka el medio ambiente circundante estaba compuesto por un mosaico de hábitats.
Las diferencias de composición entre fines del Pleistoceno, hace 30 ka, y el Holoceno pueden reflejar cambios climáticos posiblemente amplificados por la llegada del HAM. Al final del Pleistoceno Superior, la cantidad de especies asociadas a humedales, disminuye.
Según Sutikna et al (2018), hace ca 50 ka se observan cambios importantes en los conjuntos de fauna y de artefactos líticos, con extinción de H. floresiensis, Stegodon florensis insularis, Leptoptilos robustus y Trigonoceps sp. Un mayor uso de chert como materia prima alrededor de hace ca 46 ka cal es indicio de la llegada de los HAM.
Según Sutikna et al (2018), hace ca 50 ka se observan cambios importantes en los conjuntos de fauna y de artefactos líticos, con extinción de H. floresiensis, Stegodon florensis insularis, Leptoptilos robustus y Trigonoceps sp. Un mayor uso de chert como materia prima alrededor de hace ca 46 ka cal es indicio de la llegada de los HAM.
Enlaces:
- The hobbit at 10. Nature.
- Reconstrucciones tridimensionales.
- Homo floresiensis en The Knowledge Project
- Baab, K. L. (2012) Homo floresiensis: Making Sense of the Small-Bodied Hominin Fossils from Flores . Nature Education Knowledge 3(9):4
Bibliografía:
- Groves, C., and R. Wright. "Homo floresiensis: Microcephalic, pygmoid, Australopithecus or Homo?." Human Biology 57 (2006): 219-244.
- Balter, Michael. "Small but smart? Flores hominid shows signs of advanced brain." Science 307.5714 (2005): 1386-1389.
- Morwood, Michael J., and William L. Jungers. "Conclusions: implications of the Liang Bua excavations for hominin evolution and biogeography." Journal of human evolution 57.5 (2009): 640-648.
- Baab, Karen L., and Kieran P. McNulty. "Size, shape, and asymmetry in fossil hominins: the status of the LB1 cranium based on 3D morphometric analyses."Journal of Human Evolution 57.5 (2009): 608-622.
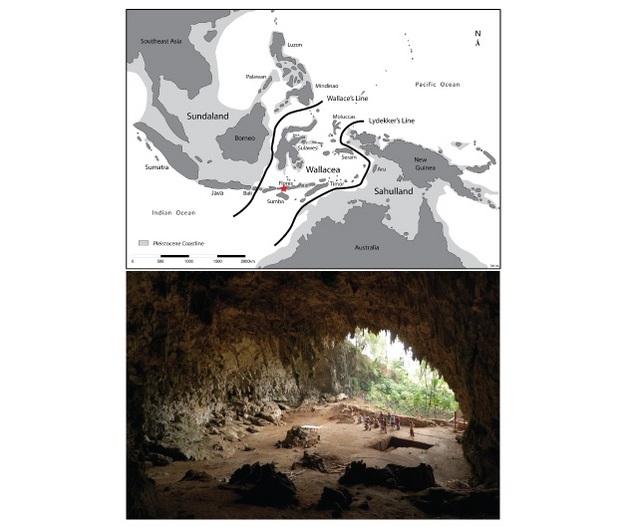 |
| Situación de la Isla de Flores y Cueva de Liang Bua |
Te puede interesar:
|