 |
| Ouranopithecus macedoniensis de Xirochori, Grecia. 10,5 Ma. Museo Nacional de Historia Natural, París. |
En lo que se refiere a Asia, los hallazgos correspondientes a este periodo están asociados a Pongo (géneros Lufengpithecus, Sivapithecus y Gigantopithecus). Gigantopithecus blacki vivió en el sur de China entre hace 2-0,3 Ma (Zhang y Harrison, 2017).
En
Europa, todavía encontramos restos de Oreopithecus
y Ouranopithecus (incertae sedis, hallado en Grecia) y en el Cáucaso un dryopitecino: el Ubadnopithecus.
Entre
8-7 Ma una crisis climática global facilitó la extensión de praderas de
gramíneas y otras herbáceas, a expensas de árboles y arbustos. En Eurasia solo
quedan Lufengpithecus, dryopitecinos
en Italia y algunas formas poco
conocidas de Extremo Oriente y Asia Menor (como Ouranopithecus turkae).
- Graecopithecus freybergi (hace 7,24-7,175 Ma). Fuss et al, 2017. Se conoce desde una única mandíbula de Pyrgos Vassilissis Amalia (Atenas, Grecia), posiblemente de un macho, hallada en 1944 por von Freyberg y descrita por von Koenigswald. En 2011, N. Spassov et al publicaron el hallazgo en Azmaka (Chirpan, Bulgaria) de un P4 superior parecido al de Ouranopithecus datado en 7 Ma.
- Las raíces mesial y distal del P4 de G . freybergi se fusionan parcialmente a aproximadamente un 47% de la longitud total de la raíz; esta condición es extremadamente rara en los grandes simios existentes pero ha sido informada en los australopitecinos y es intermedia entre S. tchadensis y Ar. kadabba. Puede representar una etapa temprana de la raíz de Tomes, que se considera diagnóstico para el clado Hominini.
- La raíz del canino es de tamaño reducido y está en el rango de las hembras de P . troglodytes y de los primeros Hominini.
Es evidente que en África
tuvieron que existir hominoideos que dieron lugar a Homini, gorilas y
chimpancés. Sin embargo la escasa existencia de fósiles correspondientes a este
periodo, oscurece el proceso.
Chororapithecus, hallado en Etiopía, datado en 8 Ma (Katoh et al, 2016) tenía la talla de un gorila y la dentición muy parecida, aunque sus molares eran de corona más baja y esmalte más grueso, lo que sugiere una dieta de vegetales fibrosos. No se puede descartar la posibilidad de una migración a África de hominoideos europeos o de Asia menor.
Carlos G. Schrago (2013) ha inferido los tamaños de población efectiva de los antepasados antropoides del linaje humano - chimpancé utilizando conjuntos de secuencias codificantes y no codificantes de unos 1.400 genes.
Según sus conclusiones, en el linaje ancestral de Anthropoidea y la de Homo y Pan se puede apreciar un patrón general de disminución del tamaño de la población. La disminución más abrupta en la población, un cuello de botella genético de unos 30.000 indiviudos durante el Mioceno tardío (12-5,5 Ma), coincide con datos biogeográficos que sugieren un evento de migración de los grandes simios desde Eurasia hacia África y la separación filogenética del ancestro de los grandes simios africanos.
Datados a finales del Mioceno Superior surgen los primeros fósiles asignados a Hominini.
 |
| Huellas de Trachilos. Gerard D. Gierliński et al, 2017. |
Bioma Pikermi
Solounias et al. (1999) señalan que muchos de los mamíferos
de la sabana africana podrían tener su origen en lo que ellos llaman el bioma
Pikermi, sobre la base de este rico yacimiento griego. Según esto, habría que
buscar el origen de los Homininae en esta parte de Europa y en Asia Menor.
En 2017, Gerard D. Gierliński et al, publicaron el hallazgo en Trachilos, oeste de Creta, de 29 huellas datadas con seguridad en ca 7,5 Ma, con un tamaño entre 94-223 mm y una forma muy similar a las huellas humanas: aquéllos individuos carecían de garras, y eran bípedos, plantígrados, pentadáctiles y con el hállux mucho más desarrollado que los otros dedos, con un cuello estrecho y una almohadilla distal asimétrica bulbosa. Los dedos se hacen progresivamente más pequeños de dentro a fuera, de modo que la región digital en su conjunto es fuertemente asimétrica. La morfología de las huellas sugiere que pertenecieron a a un miembro basal del clado Hominini, pero también podrían representar a un primate hasta ahora cuya anatomía del pie evolucionó convergentemente con la del humano.
También en 2017, Hebert Lutz, en conferencia de prensa, presentó el hallazgo en una cama del río Rhin de un diente que podría pertenecer a un hominino, con una datación de 9,7 Ma, pero la noticia fue recibida con escepticismo por la comunidad científica.
En 2005 se publicaron nuevos restos dentales y mandibulares de las localidades TM247, TM266 y TM292.
Los hallazgos de restos poscraneales, entre ellos el fémur TM266-01-63, están pendientes de publicación (Beauvilain y Watté, 2009).
El hallazgo presenta aspectos sorprendentes, por lo que ha sido clasificado a nivel de tribu como incertae sedis:
En 2017, Gerard D. Gierliński et al, publicaron el hallazgo en Trachilos, oeste de Creta, de 29 huellas datadas con seguridad en ca 7,5 Ma, con un tamaño entre 94-223 mm y una forma muy similar a las huellas humanas: aquéllos individuos carecían de garras, y eran bípedos, plantígrados, pentadáctiles y con el hállux mucho más desarrollado que los otros dedos, con un cuello estrecho y una almohadilla distal asimétrica bulbosa. Los dedos se hacen progresivamente más pequeños de dentro a fuera, de modo que la región digital en su conjunto es fuertemente asimétrica. La morfología de las huellas sugiere que pertenecieron a a un miembro basal del clado Hominini, pero también podrían representar a un primate hasta ahora cuya anatomía del pie evolucionó convergentemente con la del humano.
También en 2017, Hebert Lutz, en conferencia de prensa, presentó el hallazgo en una cama del río Rhin de un diente que podría pertenecer a un hominino, con una datación de 9,7 Ma, pero la noticia fue recibida con escepticismo por la comunidad científica.
 |
| Sahelanthropus tchadensis |
Sahelanthropus tchadensis, Brunet et al (2002)
- Holotipo: TM266-01-060-1
- Yacimientos: Toros Menalla, localidades TM247, TM266 y TM292, al oeste del desierto de Djurab, norte de Chad.
En 2005 se publicaron nuevos restos dentales y mandibulares de las localidades TM247, TM266 y TM292.
Los hallazgos de restos poscraneales, entre ellos el fémur TM266-01-63, están pendientes de publicación (Beauvilain y Watté, 2009).
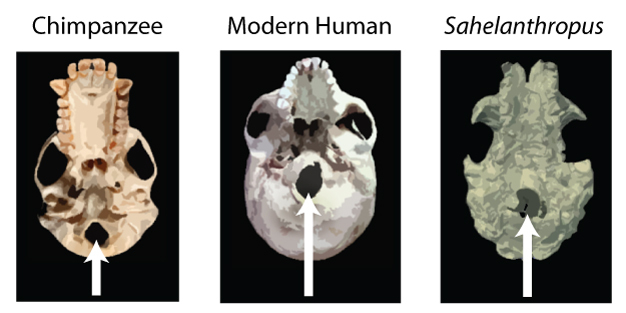 |
| El foramen magnum en el Sahelantropus se encuentra en una posición intermedia entre la del chimpancé y la del HAM (ser humano anatómicamente moderno). © 2013 Nature Education Imagen de Sahelanthropus adaptada de Zollikofer et al. 2005. |
El hallazgo presenta aspectos sorprendentes, por lo que ha sido clasificado a nivel de tribu como incertae sedis:
- Son raros los fósiles humanos hallados en aquellos parajes.
- Se localiza en el tiempo antes que los fósiles de Homini del oriente africano (considerado como foco original de los Hominini) y a 2.500 km.
- Se data en el margen de las estimaciones cronológicas establecidas por la biología molecular para la aparición de los Hominini.
- Mezcla rasgos primitivos, derivados e inusuales:
- Los arcos superciliares son muy grandes. Esta característica no aparece en los antepasados humanos hasta el Homo erectus, alrededor de 5 a 6 millones años después.
- Tambien sorprende su cara relativamente plana (débil prognatismo subnasal). Tanto los simios y los Hominini posteriores, como los australopitecos se caracterizan por su prognatismo.
- Corteza prefrontal lateral ampliada, lo que indica una reorganización cerebral incipiente, facilitada por el cambio a la posición bípeda.
RASGOS PRIMITIVOS
|
RASGOS DERIVADOS
|
OTRAS CONSIDERACIONES
| |
Capacidad craneal 378 cc.
La forma de los caninos, cónica como en chimpancés y gorilas.
Morfología de la parte posterior del cráneo similar a la de los simios.
Órbitas oculares redondeadas coronadas por un prominente torus supraorbital que recuerda el de los gorilas.
Cresta sagital en la parte posterior del cráneo. Arcada dentaria en forma de U. |
Posición del foramen magnum, caracteristica de un ser bípedo. Sin embargo, para Wolpoff (2002) esta posición es similar a la que presentan los chimpancés. Neurocráneo alargado. Reorganización cerebral. Base del cráneo larga y estrecha.
Rostro corto. Prognatismo pequeño.
Robusto reborde sobre las órbitas. Marcada constricción retroorbitaria.
Incisivos y caninos pequeños, implantados verticalmente. I1 superiores con múltiples tubérculos en la fosa lingual. Los caninos superiores no son como los de los chimpancés y muestran un extenso desgaste apical.
Premolares con dos raíces
Grosor intermedio del esmalte.
|
Vivió en las riberas de un lago. Hábitats mixtos con presencia de medios abiertos, sabanas arboladas y algunos bosques de galería. Vignaud et al, 2002. Le Fur et al, 2009. Blondel et al 2010.
Es anterior a la fecha de la separación con los chimpancés según las evidencias genéticas.
Podría ser antecesor de los chimpancés, de los humanos, de ambos o de ninguno de los dos, aunque, debido a la reorganización cerebral que muestra, es más probable que fuera antecesor de los humanos.
|
Adam van Arsdale: Sahelanthropus tchadensis
Te puede interesar:
|




