 |
| Homo habilis. Mapa conceptual. |
Homo habilis, Louis Leakey, Tobias y Napier, 1964.
Homo habilis, surge
a partir de un australopitecino grácil, sin que podamos precisar cuál. Es
posible que no lo hayamos encontrado todavía o que nunca aparezca.
Edad. 2,5 Ma (Enfriamiento del clima y desarrollo de
la sabana africana) – 1,6 Ma.
Los restos aparecieron en Olduvai a mediados de los sesenta y hasta 1964 fueron atribuidos a Australopithecus. La especie estuvo sometida a una gran polémica, que disminuyó en 1968 con el hallazgo de OH24. En 1986 se descubre OH62 con rasgos típicamente simiescos, y las dudas volvieron a surgir. En particular, se especula sobre la posibilidad de que los restos de Olduvai pertenezcan a la misma especie que los de Koobi Fora (Homo rudolfensis) o de qué forma los restos de estos dos yacimientos se pueden asociar a especies distintas.
Sigo el criterio de Alexeev y Wood de reservar habilis para los hallazgos de Olduvai.
Sigo el criterio de Alexeev y Wood de reservar habilis para los hallazgos de Olduvai.
PRINCIPALES FÓSILES
|
||
YACIMIENTOS
|
CRÁNEOS
|
POSCRÁNEOS
|
Olduvai,
Tanzania
Louis
y Mary Leakey y Philip Tobias
N.
Mbuika.
Peter
Nzube.
Donald
Johanson y Tim White
|
OH
4
OH
6
OH
7
OH
13
OH
16
OH
24 Twiggy
OH
62
OH
65
|
OH
7
OH
8
OH
35
OH
62
|
 |
| OH 7. John Reader. |
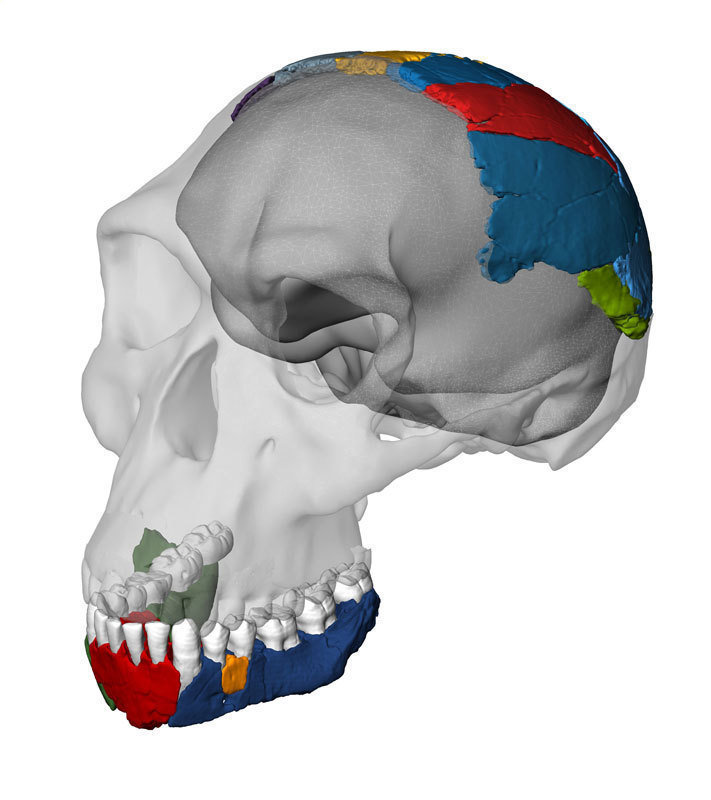 |
| Reconstrucción del cráneo de OH 7. Las partes grises están basadas en KNM-ER 1813. |
Holotipo
El holotipo de Homo habilis es OH 7 Niño de Johnny (Johnny´s child). Individuo joven hallado en 1960 por Jonathan Leakey, hijo de Louis y Mary. Huesos parietales, mandíbula provistas de 13 dientes y M3 sin erupcionar y 21 huesos de la mano que justificaron el nombre de habilis. 1,8 Ma.La mandíbula aparece distorsionada lo que ha obstaculizado los intentos de comparar este espécimen con otros fósiles.
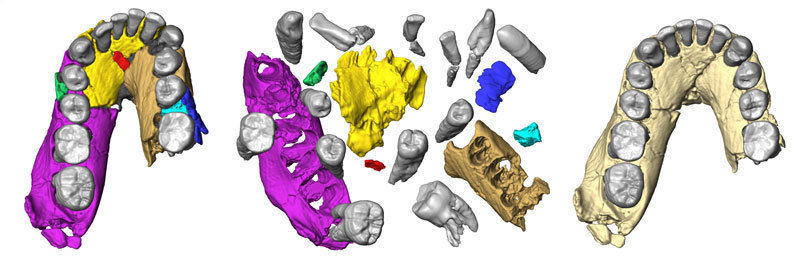 |
| Mandíbula de OH 7 tal como se ha preservado (izquierda), en partes separadas (centro) y reconstruida (derecha) |
Fred Spoor et al (2015) han reconstruido virtualmente la mandíbula y los parietales de OH 7, y los han comparado con otros fósiles de Homo temprano.
- La mandíbula reconstruida muestra un carácter muy primitivo, con una arcada dental larga y estrecha más similar a Australopithecus afarensis que a otros Homo.
- Esta morfología no es consistente con una sola especie de Homo antiguo.
- En concreto, la morfología de la mandíbula de OH 7 es incompatible con los fósiles asignados a Homo rudolfensis y con el maxilar AL 666-1. Este último es morfológicamente más derivado que el de OH 7, aunque es 500 ka años más antiguo.
- OH 7 y KNM-ER 1802 son muy similares en la forma de la arcada dental y están más cerca de OH 13 y A. afarensis que de H. sapiens y H. erectus.
- Esto sugiere que el linaje de H. habilis se originó antes de hace 2,3 Ma años.
- De la reconstrucción de los parietales, se deduce un volumen endocraneal de 729-824 cc, mayor que el obtenido en cálculos antreriores.
- La edad a la muerte se ha estimado entre 9-13,2 años, de acuerdo con los estudios radiográficos e histológicos, la longitud de las raíces de M2 y los ratios estimados de formación de esmalte y dentina en KNM-WT 15000.
Yacimientos.
Lechos I y II de Olduvai (Tanzania).
 |
| Reconstrucciones virtuales de KNM-ER 1805 (A) y OH 24 (B). En B las partes grises representan elmentos completados por espejo. Escala 5 cm. David Lordkipanidze et al, 2013. |
Paratipos.
- OH 4. Fragmentos del cráneo y dientes.
- OH 6. Fragmentos del cráneo y dientes.
- OH 8. Descrito en 1960. Dos falanges, fragmento de muela y varios huesos de un pie, completos excepto el talón y la punta de los dedos de un adulto. Su edad se estima en 1,8 Ma, con trazos mixtos entre mono y hombre, se desplazaba bípedamente. Los huesos conservan marcas de mordedura de cocodrilo (Aramendi et al, 2017; Patel et al, 2018).
- OH 13 Cinderella. Leakey, Tobias y Napier (1964). Mandíbula, dedos y fragmentos de cráneo de un adolescente, con una antigüedad de 1,6 Ma. y una capacidad craneal de 650 cc.
- OH 16, George. Leakey, Tobias y Napier (1964). Huesos de los dedos y partes muy fragmentadas del cráneo. Su edad se estima en 1,7 Ma. y su capacidad craneal en 640 cc.
- OH 24, Twiggy. 1968. 1,8 Ma. Peter Nzube. Leakey, Clarke y Leakey. Cráneo aplastado y cinco dedos. 590 cc.
- OH 35. Fragmento distal de tibia. Presenta marcas de mordedura que no caen en el rango de las producidas por cocodrilo (Aramendi et al, 2017). Según Marchi et al (2019), la fuerza relativa fibular/tibial se aproxima a la de los grandes simios, lo que indica vida arbórea y, en particular, la capacidad de escalada vertical
- OH 62. Dik-dik hominid. 1986. 1,8 Ma. Tim White. Johanson et al (1987). Incluye un paladar, fragmentos del cráneo, restos del húmero y del radio, parte del fémur y de la tibia y otros restos menores. En la zona del paladar y la dentición se parece a OH 24, ER 1813 y ER 1470 más que a los australopitecinos. Sin embargo, las medidas de las partes superior e inferior del cuerpo son similares a las de los australopitecinos. Medía 1,05 m y sus brazos son muy largos en comparación a las piernas, con rasgos parecidos a Lucy. Adscrito al habilis a partir de su mandíbula y dentición.
- OH 65. 1995. Olduvai. 1,8 Ma. Blumenschine et al (2003). Clarke (2012). Mandíbula superior completa y parte inferior de la cara. Datado en 1,8 Ma. Por sus similitudes con ER 1470, se ha sugerido que OH 65 puede reconducir a una reclasificación de los habilis. La dirección de las estriaciones dentales sugiere que perteneció a un individuo diestro lo que constituye la evidencia más temprana de lateralización (Frayer et al, 2016).
 |
| OH24. Homo habilis. Smithsonian. |
Morfología
- Capacidad craneal 610 cc (mínimo 510; máximo 824). Coeficiente de encefalización próximo a 3.
- Hay un aumento de la corteza cerebral, en las áreas asociativas parietales (área de Wernicke) y occipitales, sin correspondencia con el aumento corporal, obligando a producir reestructuraciones corticales importantes. El lóbulo frontal inferior (área de Broca) aumenta en superficie y complejidad.
- Mayor irrigación sanguínea del cerebro.
- Cráneo más grácil: menor desarrollo de crestas musculares, anchura y altura facial más reducida, pilares faciales más ligeros.
- El hemisferio derecho es ligeramente mayor que el izquierdo y se extiende más hacia el polo frontal mientras que el izquierdo lo hace hacia el polo occipital.
- El aumento cerebral pudo deberse a una mutación en los genes reguladores (heterocronía). El aumento cerebral produjo una mayor inmadurez neurológica en los recién nacidos, alargando el tiempo de infancia. Esto condujo a una conducta social protectora.
- Huesos de la bóveda craneal delgados.
- Curvatura del hueso parietal presente en el perfil de la caja craneana mientras que en los Australopithecus está muy poco afianzada.
- El ángulo formado por las dos partes del hueso occipital es abierto (afilado en los australopitecos.
- Ausencia de cresta sagital. Huellas acusadas de las inserciones musculares en el cráneo.
- Foramen magnum en posición adelantada.
- Toro supraorbital.
- Escaso prognatismo. La región del mentón está retrasada. Aunque OH 7 y KNM-ER 1802 son muy similares en la forma de la arcada dental y están más cerca de OH 13 y A. afarensis que de H. sapiens y H. erectus.
- Huesos nasales que se ensanchan inferiormente. Rebordes de la abertura nasal verticales y estrechos.
- Porción media de la cara relativamente estrecha, con región malar casi vertical, más desarrollada que la parte inferior.
- Dentición.
- Tercer molar apenas más grande que el segundo.
- Del examen de los dientes de deduce un patrón de desarrollo rápido, como el de los chimpancés.
- Esmalte grueso.
- Tendencia hacia el estrechamiento bucolingual y al alargamiento mesiodistal, sobre todo en los premolares inferiores y los molares.
- Un metro de estatura. Peso corporal de 50 kg para los machos y 30 kg para las hembras, lo que implica un dimorfismo sexual acusado.
- Clavícula semejante a la de los humanos modernos. Los restos de hombros son muy escasos y sugieren que su configuración era similar a la de los homínidos tempranos.
- Extremidades inferiores más largas relativamente que las de los australopitecos. Pudiera ser que H. habilis retuviese las extremidades inferiores de un trepador (Groves, Stringer, Walker, R. Leakey). Últimas falanges de las manos anchas. La articulación entre pulgar y metacarpo se parece a la de sapiens. Las marcas de unión de los tendones de los músculos flexores son fuertes y las falanges son curvadas.
- OH 7 posee falanges proximales y medias con marcada curvatura palmar, diáfisis mediolateral ampliada y fuertes flexores, lo que se asocia comúnmente con la locomoción arbórea. Además, la falange distal pollical de OH 7 también carece de un lugar de inserción para el tendón flexor largo y otras características que se relacionan con el agarre de precisión (Moyà-Solà et al, 2008). Por tanto, las falanges de OH 7 no encajan en un patrón Homo.
- Bipedestación completa.
- Pie relativamente más corto como en los humanos modernos.
- Huesos del pie parecidos a los de humanos modernos, con un hallux corto y robusto, adducido.
- La articulación del tobillo difiere un tanto y el tercer metatarso parece robusto.
- No ha podido ser confirmada la presencia del arco longitudinal.
- Astrágalo diferente al patrón humano.
 |
| OH 86. De izquierda a derecha, vistas dorsal, lateral, palmar (la parte distal arriba en todos los casos) y proximal. Barra de escala, 1 cm. Domínguez-Rodrigo et al, 2015. |
OH 86 y un morfo adicional en Olduvai
OH 86 es una quinta falange proximal izquierda completa recientemente hallada en Philip Tobias Korongo (PTK) en la Garganta de Olduvai (Tanzania), datada en más de hace 1,84 Ma, asociada a fauna y abundantes artefactos de piedra.
- Esta hipótesis es apoyada tanto por criterios cualitativos, incluyendo la asimetría de las crestas de los flexores y la tróclea distal, así como la orientación de esta última y el contorno palmar de la base, como por criterios cuantitativos y por el análisis discriminante.
- Atendiendo al tamaño de OH 86, los individuos de este morfo disfrutarían de un tamaño mucho mayor al conocido para Homo habilis, quizá en la línea evolutiva de Homo erectus.
Discusión filogenética.
- ¿Homo habilis es el ancestro de erectus?
- El hallazgo en Turkana de KNM-WT 15000, Turkana Boy datado en 1,6 Ma implica que habilis y erectus convivieron en la misma área durante medio millón de años y convierte en insostenible la secuencia habilis->erectus. OH 62 se ha datado en 1,8 Ma.
- Para Conroy (2005) la solución es negar que OH62 pertenezca a habilis.
- Para Arsuaga (2006) es posible que erectus descienda de una población de habilis mientras otras poblaciones permanecieron en estasis.
- OH 86 apoya la existencia de un morfo distinto a habilis quizá en la línea evolutiva de Homo erectus.
- ¿Por qué ubicar los restos de Olduvai en Homo y no en Australopithecus?
- La taxonomía toma en cuenta, por encima de cualquier otra consideración, la morfología de los especímenes y en esto se basó Leakey (rasgos craneodentales), pero estas diferencias con los australopitecinos no están claras. Con el tiempo, se ha impuesto también una identificación basada la conducta. Las diferencias entre Australopithecus y Homo son básicamente culturales:
- la construcción de herramientas en piedra para el carroñeo, basada en la presencia de un cerebro lo bastante grande para llevar a cabo las complejas operaciones cognitivas necesarias. Estamos ante el primer homínido que fue capaza de modificar intencionalmente el entorno. Homo sería el responsable de la cultura lítica olduvaiense y H. habilis su primer representante. Habilis significa en latín: capaz, habilidoso, mentalmente desarrollado, vigoroso.
- Cambios conductuales. Los yacimientos parecen indicar que, tras el rápido troceamiento de animales muertos con herramientas preparadas, trasladaban el botín obtenido a lugares específicos de reunión del grupo, donde se repartía. Compartir los alimentos y proteger las crías son las innovaciones conductuales más destacadas.
- Gracias a las herramientas, el índice de eficacia en la adquisición de nutrientes es más elevado, lo que permite costear un cerebro más grande.
- Esta interpretación está oscurecida por el hallazgo de un morfo adicional representado por OH 86, que añade incertidumbre a la autoría de las herramientas.
- Mark Collard y Bernard Wood (1999) propusieron la categoría de Australopithecus habilis, basándose en en que su ontogenia es comparable a la de los australopitecinos, mientras que el crecimiento de Homo se extiende durante 18 años, con una infancia prolongada y una larga adolescencia.
 |
| Problemas de clasificación de Homo habilis |
Modos de Vida.
- Construcción de herramientas. (Cultura Olduvaiense (Modo I): Choppers
(piedras cortadoras labradas por una sola cara quizá de un solo golpe preciso
sobre la piedra núcleo) y lascas).
- Es probable una dieta con mayor contenido de carne. Omnívoro
oportunista. ¿Carroñero?¿Cazador de piezas pequeñas? ¿Ambas cosas? Según
Henry Bunn, en un yacimiento de la garganta de Olduvai se encontraron
evidencias de caza con emboscada hace 1,6 Ma.
- Machacaban huesos para obtener el tuétano.
- Elaboración de complejas funciones cognitivas para la actuación en grupo
y la enseñanza de tecnología.
Adam van Arsdale
Enlaces:
Gómez
Castanedo, A: (2012), "Los orígenes del género Homo. Una exploración de la
innovación como motor evolutivo". Revista Nivel Cero, 13. Santander.
Bibliografía:
- Blumenschine, Robert J., Ian G. Stanistreet, and Fidelis T. Masao. "Olduvai Gorge and the Olduvai Landscape Paleoanthropology Project." Journal of Human Evolution 63.2 (2012): 247-250.
- Tobias, Phillip V. "Encore Olduvai." Science 299.5610 (2003): 1193-1194
Te puede interesar:
|
No hay comentarios:
Publicar un comentario