.png) |
| Australopithecus africanus. Mapa conceptual. Una datación de STW 573 atrasa la presencia de la especia a hace 3,67 Ma, pero no todos los investigadores aceptan esta fecha. |
Australopithecus africanus, Dart 1925.
- Holotipo: Taung 1. Niño de Taung.
- Yacimientos: Todos ellos en Sudáfrica.
- Makapansgat
- Sterkfontein
- Gladysvale
- Taung
3,67-2,3 Ma. La datación es muy problemática. Lo más probable es que sea un poco anterior a 2,5 Ma.
Su nombre significa mono
del sur de África.
Dart propuso el género Australopithecus y la familia Homo-Simidae para los fósiles de la cantera de Taung, recuperados en 1924. Ante la falta de grandes caninos, consideró que estos hominini habrían sido cazadores despiadados utilizando una industria osteodontoquerática. Actualmente se considera que el africanus no fue un depredador sino una presa.
En 1936 se descubrieron en Skertfontein los restos de TM 1511 y TM 1512 y en 1947 los de STS 5.
Dart propuso el género Australopithecus y la familia Homo-Simidae para los fósiles de la cantera de Taung, recuperados en 1924. Ante la falta de grandes caninos, consideró que estos hominini habrían sido cazadores despiadados utilizando una industria osteodontoquerática. Actualmente se considera que el africanus no fue un depredador sino una presa.
En 1936 se descubrieron en Skertfontein los restos de TM 1511 y TM 1512 y en 1947 los de STS 5.
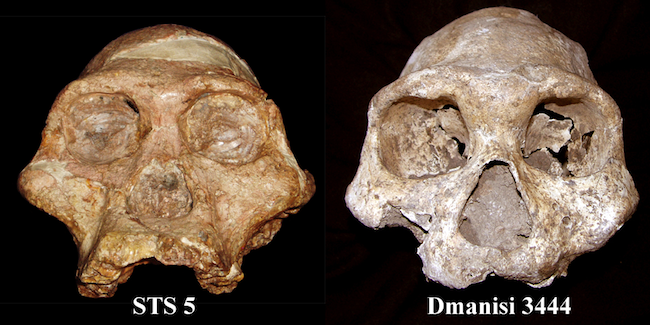 |
| Comparación entre STS 5 y Dmanisi 3444 |
Hallazgos
La acumulación de miles de huesos de diversas especies en
estas cuevas puede deberse a caídas, arrastres o comportamientos de acumulación
de algún depredador, incluyendo aves.
- Limeworks Cave miembros 3 y 4, Makapansgat, Northern Province, Sudáfrica. 3-2,6 Ma. Desde 1946. 27 especímenes de al menos 10 individuos. Mosaico de hábitats. El estudio de isótopos estables refleja un consumo de hierbas y juncos (C4) (Sponheimer y Lee-Thorp, 1999).
- MLD 1. Calvaria parcial de adulto.
- MLD 2. Mandíbula.
- MLD 6/23. Mandíbula.
- MLD 9. Mandíbula.
- MLD 11/30. Maxilar y dientes asociados. Edad al morir, 3,42 años (Smith et al, 2015).
- MLD 18. Mandíbula.
- MLD 37/38. Calvaria parcial de adulto.
- MLD 40. Mandíbula.
 |
| MLD 2 |
 |
| MLD 18 |
- Taung, North-West Province, Sudáfrica. 3-2,6 Ma. 1924. Mosaico de habitats.
- Taung 1. El niño de Taung (Taung child). 1924. Raymond Dart. Parte del cráneo (molde craneal), cara y mandíbula de un individuo joven (3-4 años), con la dentición decidua (Kuhn et al, 2016). El primer molar permanente empieza a emerger. Con dificultades para una buena datación. Su capacidad craneana es de 410 cc y se supone que de adulto tendría una capacidad de 440 cc. El gran tamaño del cerebro y pequeño tamaño de los caninos, junto con la posición centrada del foramen mágnum convencieron a Dart de su bipedia. Philip Tobias propuso que procedía de una cueva (McKee, 2015; Jeffrey K. McKee y Kevin L. Kuykendall (2016); por el contrario Philip J. Hopley et al, 2013 consideran que procede de la superficie del paleosuelo). Presenta otros rasgos llamativos:
- Abertura nasal pequeña.
- Frente muy vertical.
- Zona supraorbital poco pronunciada.
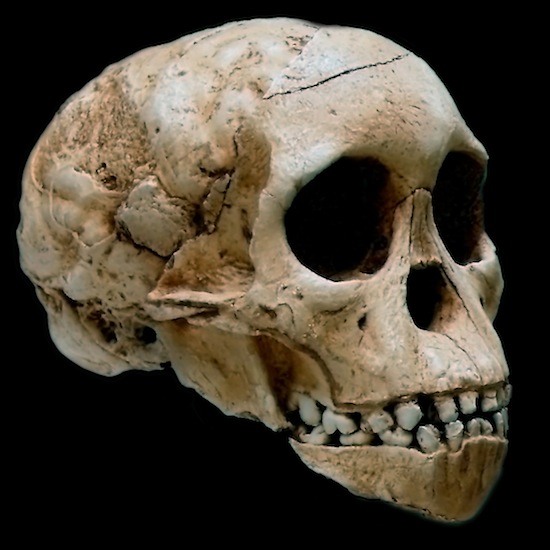 |
| Taung 1 |
- Sterkfontein, Gauteng, Sudáfrica. 2,6-2 Ma. Mosaico de hábitats. Diversas proporciones de C3 y C4 en la dieta, lo que indica una mezcla de tipos de vegetación y posiblemente termitas. De aquí se ha extraído la muestra más extensa.
- STS 1. Palatal.
- STS 2. Maxilar y dientes asociados. Edad al morir, 2,52 años (Smith et al, 2015).
- STS 5. La Señora Ples (Mrs. Ples). 1947. 2 Ma. Robert Broom. Se trata de un cráneo muy bien conservado correspondiente a una hembra madura (B. Villmoare et al, 2013). Su capacidad es de 485 cc.
- STS 14. 1947. 2,5 Ma. Robert Broom y J.T. Robinson. Es el mejor especimen de africanus recuperado hasta el momento. Una casi completa columna vertebral con vértebras lumbares (seis, mientras que el HAM tiene cinco y el chimpancé tres) y torácicas, pelvis, fragmentos de costillas y una parte de fémur. Por sus pequeñas dimensiones en principio se atribuyó a una hembra pero actualmente se estima es de un macho. Recientemente se ha especulado que Sts 5 y Sts 14 corresponden al mismo individuo. Fuertes evidencias de bipedación, aunque la zona lumbar es de pequeño tamaño. El individuo era delgado y con una estatura de 122-137 cm (Robinson, 1972).
- STS 17.
- STS 24. Maxilar y dientes mandibulares asociados. Edad al morir, 4,35 años (Smith et al, 2015).
- STS 71. Cráneo. McNulty (2012) encuentra evidencias de pedomorfismo.
- STW 13.
- STW 89. Segundo metatarsiano.
- STW 183. Maxilar.
- STW 252. Cráneo.
- STW 352. Calcáneo. Zeininger et al, 2016.
- STW 384. Mandíbula.
- STW 431. Esqueleto parcial. Incluye seis vértebras lumbares. Odes et al, 2017.
- STW 432. Esqueleto parcial de tamaño grande.
- STW 498. Maxilar y mandíbula.
- STW 505. Cráneo. 500 cc.
{kind=link}
 |
| Modelo 3D del hallazgo de Little Foot. |
 |
| STW 573. Little Foot. |
- STW 573. Little Foot. 1994-1997. Gruta Silberberg. 3,67 Ma. (Clarke, 2008; Kramerks y Dirks, 2016).
- En 1994, Ron Clarke reconoció cuatro huesos del pie izquierdo de un hominino en dos cajas de restos de fauna procedentes de una cantera de Sterkfontein. En 1997, Clarke, Stephen Motsumi y Nkwane Molefe lograron localizar más huesos de ese pie incrustados en la roca. Su tamaño justificó el apodo de Little Foot. Durante los siguientes 15 años, se recuperaron fragmentos de roca con huesos incrustados, mediante un trabajo delicado con pequeños taladros. En los cinco años siguientes, los fósiles fueron limpiados y reconstruidos. El resultado es el espectacular esqueleto identificado como Stw 573, que conserva incluso algunas partes articuladas (brazo izquierdo y mano) y el cráneo completo.
- El esqueleto perteneció a una hembra entre 1,20-1,30 m de altura. Muestra una combinación de vida terrestre y arbórea. STW 573 es el primer esqueleto de hominini del que se pueden deducir las proporciones de las extremidades inequívocamente. Demuestra que algunos homininos tempranos, en el momento de la formación de las huellas de Laetoli (hace 3,6 Ma), poseían un cuerpo grande, con extremidades posteriores más largas que las extremidades anteriores. Los estudios de modelación de primates existentes indican que el índice intermembral (IMI) de STW 573, es bajo para un gran simio no humano, y habría mejorado sustancialmente la economía de la bipedación en distancias de medias a largas, perjudicando la escalada, pero que todavía era demasiado alto para un desplazamiento eficaz con carga (Robin Huw Crompton et al, 2018).
- Las extremidades inferiores muestran muchas características modernas.
- El pie izquierdo conserva astrágalo, escafoides articulado con la cabeza del astrágalo, primer cuneiforme articulado con el escafoides y mitad proximal del dedo gordo articulado con el cuneiforme (Clarke y Tobias, 1995), con un mosaico de rasgos humanos y simiescos que sugiere una bipedación incompleta y un entorno forestal.
- Mano muy grande. Observaciones preliminares de la morfología de la mano, describen una articulación trapecio primer metacarpiano diferente a la de los seres humanos y los chimpancés, un pulgar humano robusto y falanges proximales tan curvadas como las de Au. afarensis. No hay inferencias acerca de su capacidad manipuladora, pero las falanges curvadas sugieren el uso de la mano para la locomoción y la ausencia de herramientas de piedra en el miembro 2 de Sterkfontein puede indicar que la evolución de las capacidades de manipulación pudo estar relacionada con el uso de herramientas no líticas (Kivell, 2015).
- Brazos largos.
- Cara plana, frente amplia. Dentición de gran tamaño. Clarke y Kuman, 2018.
- El cráneo incluye la primera vértebra cervical, parecida a la de los chimpancés, lo que indica un movimiento de la cabeza adaptado a la trepa. Beaudet el al, 2020.
- Datación:
- La datación de las capas por encima y por debajo de los fósiles, mediante técnicas de desintegración de isótopos U-Pb (uranio-plomo), dio un resultado de alrededor de 2,2 Ma, menor del esperado si se consideran la fauna asociada y la posición estratigráfica. Estudios estratigráficos, micromorfológicos y geoquímicos recientes revelaron que la estratigrafía es mucho más complicada de lo que se pensaba, con procesos post-deposicionales localizados que condujeron a la creación de huecos en las capas donde se asentaba el esqueleto, que se fueron rellenando a lo largo de miles de años (espeleotemas). Según Laurent Bruxelles et al (2014) las fechas propuestas en base a la deposición de coladas pueden dar sólo una edad mínima para Stw 573, ya que se formaron mucho después. Para los autores, es probable que la fecha se acerque a los 3 Ma.
- Darryl E. Granger et al (2015) han demostrado que la brecha que contiene a STW 573 no sufrió reelaboración significativa, y fue depositada hace 3,67 ± 0,16 Ma.
- Según Kramers y Dirks (2017) las muestras tomadas cerca del lugar de hallazgo de StW 573 imponen una edad máxima para el fósil de 2,8 Ma. Los autores proponen un escenario de enterramiento de dos etapas.
- El equipo de Clarke ha publicado el resultado de un nuevo estudio que confirma sus estimaciones anteriores de 3,67 Ma (Bruxelles et al, 2018).
- Según Travis Rayne Pickering, Ron J. Clarke y Jason L. Heaton (2014), los restos del Miembro 2 de Sterkfontein corresponden a individuos que entraron en la cueva por su cuenta, ya sea por caída o por entrada intencional, y no pudieron salir. El paleoambiente se caracterizó por colinas de roquedales cubiertas por matorrales. Además, el fondo del valle pudo haber retenido agua permanente durante todo el año.
- STW 576. Palatal adulto con dentición completa, único caso disponible para A. Africanus (Clarke y Kuman, 2018).
- STW 578. Cráneo parcial. Beaudet et al, 2018. Los sedimentos de donde procede, se han datado en hace 4 Ma, y por tanto constituiría el ejemplar de australopitecino más antiguo que se conoce en los sistemas kársticos de Sudáfrica.
- STW 670. Fragmento proximal de una primera costilla del lado derecho. Tawane et al, 2016.
 |
| TM 1511. Créditos: Jason Heaton. |
- TM 1511. 1936.
- TM 1512. 1936. Robert Broom Se trata de partes de la cara, mandíbula inferior y bóveda del cráneo.
- Gladysvale, 13 km al nordeste de Sterkfontein. Datados entre 2,4-2 Ma.
 |
| Ángulos de prognatismo de Sts 5 (A), Sts 17 (B), y Sts 71 (C). Las imágenes no están a escala. B. Villmoare et al (2013). |
HALLAZGOS CRANEALES
|
|||
Cráneos completos
|
Cráneos fragmentarios
|
Mandíbulas
|
|
STERKFONTEIN
2,6-2 Ma
|
STS 5
|
STS
17
STS
52a
STS
53
STS
63
STS
71
STW
13
STW
73
STW
252
STW
505
|
STS
7
STS
36
STS
52b
STW
384
STW
404
STW
498
STW
513
|
TAUNG
2,6 Ma
|
TAUNG 1 (juvenil)
|
||
MAKAPANGSGAT
3-2,6 Ma
|
MLD 6
MLD 9
MLD 23
|
MLD
2
MLD
12
MLD
22
MLD
29
MLD
34
MLD
40
MLD
45
|
|
Morfología.
- Un cráneo más alto y corto, con cresta sagital rara y cresta temponucal no compuesta. La base del cráneo es más estrecha en relación con su longitud y el foramen magnum se encuentra más posteriormente. El plano occipital del hueso occipital es más alto y el plano de la nuca es más llano y más horizontal.
- El torus supraorbitario adelgaza lateralmente de arriba abajo y ocasionalmente se divide en distintos componentes supraorbitario y superciliar (Lockwood y Tobias, 1999).
- La fosa de la mandíbula es más profunda, en promedio, con una eminencia articular fuerte.
- El elemento timpánico está más orientado verticalmente, por lo general tiene una cresta petrosa distinguible y se estrecha medialmente a una proyección de Eustaquio diferente.
- El drenaje venoso del endocráneo se realiza predominantemente a través del sistema transverso-sigmoideo.
- La abertura nasal está rodeada de prominentes pilares anteriores, incluso en individuos jóvenes, y la placa subnasal es plana a ligeramente convexa y sagital, mucho menos proyectada en relación al eje bicanino
- Dentro de la cavidad nasal, la bajada al piso nasal y la inserción vomeral anterior, se produce inmediatamente posterior al nasoespinal, por lo general sin una plataforma de intervención.
- El hueso cigomático presenta un patrón variable de prominencia en la transición a la apófisis temporal y una fuerte inflexión sagital a través de la apófisis frontal / transición de la superficie facial, que se combinan para crear un hueco central facial en algunos individuos.
- Las raíces de la apófisis cigomática por lo general se originan más anteriormente (por encima de P4/M1 P4/P3) con un margen inferior recto, superolateralmente divergente.
- El paladar, en promedio, más profundo, con un segmento premaxilar con la parte inferior flexionada y filas de dientes divergentes posteriormente.
- El corpus de la mandíbula es más robusto (amplitud en relación a profundidad en M1) y tiene una mayor superficie lateral por debajo de los premolares y una sínfisis con un perfil más vertical y recto. Ramus alto. Carencia de barbilla.
- Dentición.
- Incisivos pequeños y anchos.
- Caninos poco proyectados.
- El desgaste canino es exclusivamente apical. Diastemas C/P3 e I2/C son menos frecuentes.
- Poscaninos más largos y sobre todo anchos con los ápices de las cúspides molares situados centralmente en las coronas en algunos individuos. La dentición crece progresivamente, siendo más pequeña la anterior.
- Premolares de tamaño intermedio. El P3 es uniformemente bicúspide
- El gradiente en adultos de desgaste oclusal anterior-posterior es más débil.
- El análisis de elementos traza revela ha revelado que los bebés de A. africanus consumieron predominantemente leche materna durante el primer año después del nacimiento. La acumulación cíclica de litio sugiere recursos fluctuantes y adaptaciones fisiológicas a esta inestabilidad, con ingesta de leche en momentos de desafío nutricional, como se observa en los orangutanes salvajes modernos (Joannes-Boyau et al, 2019).
- Esqueleto poscraneal muy similar (White, 2002), aunque con algunas diferencias en las proporciones de las extremidades superiores e inferiores. Los rasgos sugieren bipedación (Grine, 2000). El ángulo bicondiliar refleja que las rodillas y los pies estuvieron muy próximos al caminar.
- Meir M. Barak et al (2013), basándose en la orientación de la articulación del tobillo durante la carga en la marcha y la orientación principal de los puntales trabeculares en la tibia distal cuya función es soportar las fuerzas de compresión, en STW 358 y STW 389, del miembro 4 de Sterkfontein, consideran que estos hominini cargaban principalmente sus tobillos en una postura relativamente extendida, como los seres humanos modernos, y diferente a la de los chimpancés. Su bipedación por tanto sería eficiente, con las extremidades extendidas.
- No hay unanimidad con respecto a las proporciones físicas del taxón. Reed et al (1993) proponen un dimorfismo sexual similar. Algunas estimaciones apuntan a unas proporciones similares a las de los HAM mientras que otras están más cerca del patrón simiesco.
- Los parásitos Pediculus humanus (piojo) y Pthirus pubis (ladilla) son especies no relacionadas específicas para el humano, el primero en la cabeza y el segundo en el pubis y esta coexistencia solo es posible gracias a que estas dos zonas están separadas por un amplio espacio lampiño. Otros mamíferos muestra un solo parásito homologable. De acuerdo con los datos genéticos, la primera especie tuvo origen hace 7 Ma y la segunda hace 3 Ma, momento en que abundaba Australopithecus africanus, lo que sugiere que era lampiño.
Características de STS 5 “la señora Ples”
- 1,38 – 1,15 m. 41 – 30 kg.
- Cráneo redondeado, con un frontal más recto y un occipital más
convexo. Caja craneal estrecha, de unos 485 cm3. Asimetría
cerebral más acusada, al igual que la densidad de la red meníngea media.
El rango de variación de todos los ejemplares hallados está entre 435-560
cc.
- Arco supraorbital no muy desarrollado, forma un reborde que precede
a la placa del frontal, glabela algo pronunciada, cresta occipital débil y
plano nucal bajo y llano.
- Base del cráneo más arqueada y corta. Foramen mágnum centrado (bipedestación)
y región del mastoideo pequeña.
- Caracteres asociados al desarrollo de un aparato masticador cada
vez más potente.
- Paladar profundo en la parte anterior. Pilares a ambos lados del
orificio nasal. El clivus naseoalveolar es plano y el umbral entre estas
dos partes está apenas sugerido. Raíz anterior del arco cigomático encima
de P4, más adelantada que en afarensis.
- Rostro menos prognato. La mandíbula tiene un ramus alto y carece de
barbilla. Sínfisis mandibular menos inclinada y torus transversales menos
desarrollados. Las ramas horizontales de la mandíbula son robustas.
Desarrollo más marcado de la cara anterior del músculo temporal y de la
constricción posorbital.
- El tamaño de los dientes incrementándose de delante hacia atrás.
Superficie trituradora importante. Esmalte grueso. Incisivos anchos y
pequeños, con raíces rectas. Caninos reducidos que se desgastan a partir
de la punta. Premolares relativamente grandes, con coronas de dos
cúspides. Molares más desarrollados bucolingualmente.
- Estudiando los dientes del Niño
de Taung, Holly Smith por un lado y Timothy Bromage por otro llegaron
a la conclusión de que Australopithecus
tenía un desarrollo rápido, como el de los chimpancés.
- Mano similar a la humana (Skinner et al, 2015; pero Amécija et al, 2015; réplica Skinner et al, 2015).
- Las caderas y miembros posteriores indican bipedia no del todo
desarrollada (se ha hallado una pelvis completa, STS-14). Atrágalo
bastante humano mientras que tarso y primer metatarsiano son similares al aferensis. El tarso de STW 573 (Little Foot) parece más derivado.
- Constitución grácil.
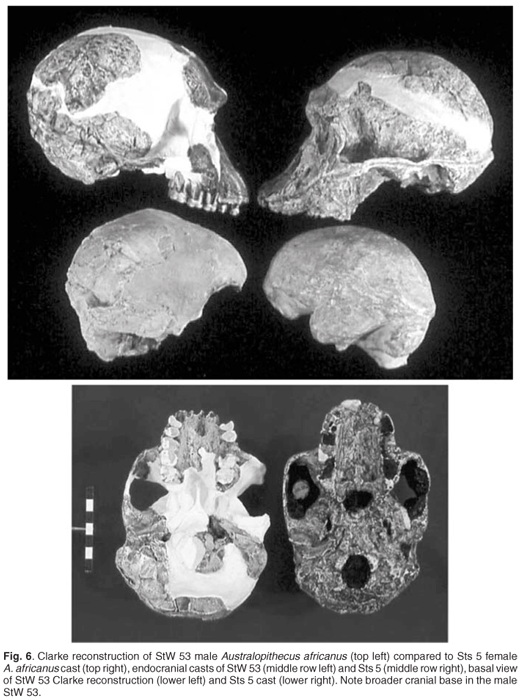 |
| Reconstrucciones de STW 53 y STS 5. Clarke. |
Discusión filogenética
Pese a la abundancia de restos, la condición del A. africanus ha sido muy debatida, quizá por la dificultad de
relacionar esta especie en términos filogenéticos con las posteriores (Paranthropus robustus y Homo habilis). Los yacimientos
sudafricanos son de difícil extracción y datación.
Los rasgos craneodentales se aproximan más a Homo que a afarensis, mientras que las proporciones físicas parecen más primitivas que las de afarensis. Las uniones de los huesos de los brazos son también más arcaicas.
Los rasgos craneodentales se aproximan más a Homo que a afarensis, mientras que las proporciones físicas parecen más primitivas que las de afarensis. Las uniones de los huesos de los brazos son también más arcaicas.
- Para Lockwood y Tobias A. africanus es la especie ancestral tanto de Paranthropus como de Homo. La gracilidad de A. africanus se ha establecido por comparación con el Paranthropus, pero de todas formas es el posible iniciador del linaje robusto.
- White et al (1981) y Rak (1985) lo excluyen como ancestro de Homo porque consideran que está en la línea de especialización dental de los robustos. Cela y Ayala (2001) lo sitúan como el posible iniciador del linaje robusto.
- Según Brian Willmoare y William Kimbel (2011), los pilares anteriores del rostro del A. africanus son huecos mientras que los de P. robustus y P. boisei contienen un tejido denso y esponjoso. De ello deducen que los parántropos no están relacionados filogenéticamente con A. africanus.
- Debido a que su dentición parece más moderna que la de afarensis, Tobias, Clarke y Berger proponen que los primeros Homo están relacionados con africanus y no con afarensis.
- Berger y Hilton-Barber, 2001, creen que africanus podría descender de anamensis o de algún taxón similar que se expandió por África del Este y del Sur y dio lugar a afarensis en entornos más abiertos y a africanus en entornos más cerrados.
- Kimbel et al (2006), sugieren que africanus evolucionó a partir de una población similar a la de Hadar.
- Para Walker et al (1986) africanus desciende de afarensis pero a través de una población intermedia.
- Esta opinión no es compatible con la reciente datación de STW 573 que convierte a africanus en contemporáneo de afarensis (Darryl E. Granger et al, 2015).
Inferencias sobre sus modos de vida.
Según Sandi Copeland (2011), tras estudiar la procedencia de
los individuos a partir de rastros en los dientes, los grupos de africanus practicaban la filopatría (los
machos se unían a hembras de fuera).
Para Ferrán Estebaranz Sánchez et al, A. africanus presenta un patrón de microestriación muy homogéneo y
en gran medida semejante al de chimpancés y gorilas, por lo que su dieta podría
representar el modelo ecológico ancestral de los hominoideos, una dieta
generalista de ambientes cerrados (hojas y frutos frescos) con incorporación de
juncos y hierbas de tipo C4 y pequeños invertebrados en la estación seca. La ausencia
de herramientas líticas, una flora con lianas y una fauna con monos y sin herbívoros
hace pensar que viviera en un ambiente boscoso en el límite temporal con la formación
de la sabana africana.
Según Wynn, Cerling y Sponheimer (2013), su dieta estaba compuesta de alimentos procedentes de árboles y arbustos, pero también de hierbas y juncias, con gran variación entre los individuos.
Según Wynn, Cerling y Sponheimer (2013), su dieta estaba compuesta de alimentos procedentes de árboles y arbustos, pero también de hierbas y juncias, con gran variación entre los individuos.
Era bípedo o al menos caminaba erguido.
Australopithecus prometheus
Dart asignó este nombre a los primeros Hominini descubiertos en Makapansgat, que pronto quedaron incluidos en Australopithecus africanus. El nombre deriva de los restos de animales asociados, que Dart creyó quemados.
Ronald Clarke defiende actualmente la idea de dos especies en los yacimientos de Makapansgat y Skertfontein. Asigna a una segunda especie los especímenes siguientes:
- MLD 2
- STS 1.
- STS 71.
- STW 183.
- STW 252. Mayer, 2016. Stw 252, Varios fragmentos de cráneo y dentadura completa de un individuo de 7-9 años de edad, dado que los M3 están sin erupcionar completamente y por la ausencia de desgaste. Hallado en 1984 por Alun Hughes, y descrito por Clarke en 1988.
- STW 384.
- STW 431.
- STW 498.
- STW 505. Cráneo.
- STW 573.
- Morfología muy parecida a los elementos disponibles de Australopithecus anamensis (por ejemplo, las superficies articulares tibial y humeral proximales y distales) y también existen similitudes con las características de la escápula, en particular, de KSD-VP-1/1 (A. afarensis de Woranso-Mille). Sin embargo, las similitudes más cercanas son con el esqueleto parcial STW 431 del miembro 4. de Sterkfontein. Cuando se consideran juntos, tanto el STW 573 como el STW 431, expresan una morfología de la articulación de la cadera bastante distinta de la de Sts14 (Australopithecus africanus), y un fémur proximal de un presunto A. africanus de la Caverna Jacovec en Sterkfontein, STW 598 (Robin Huw Crompton et al, 2018).
- El cráneo es similar a A. prometheus y diferente a A. africanus. Una de sus características más interesantes es un patrón de desgaste dental anterior muy pesado, a diferencia del que se encuentra en A. africanus, pero que se asemeja al encontrado en A. anamensis en 4.17 Ma (Clarke y Kuman, 2018).
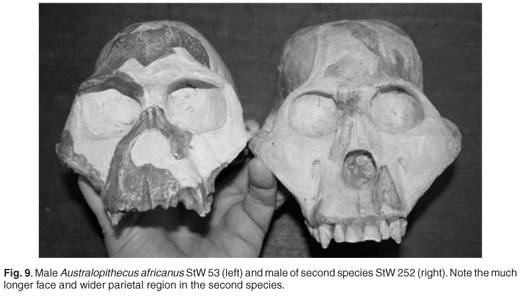 |
| STW 53 y STW 252. Clarke. |
Clarke (2008) define esta segunda especie por las siguientes características:
- Molares y premolares grandes y bulbosos, con cúspides situadas hacia el centro de la corona.
- Caninos e incisivos grandes en relación a los dientes posteriores.
- Pómulos prominentes, de manera que en una vista lateral, la región nasal se oculta detrás de los pómulos.
- A partir de una posición a mitad de camino hasta el borde lateral de la abertura nasal, el proceso frontal del maxilar se extiende lateralmente en lugar de curvado hacia atrás, como lo hace en A. africanus.
- La región interorbital es amplia, mientras que en A. africanus es estrecha.
- Nasion situado encima de la sutura frontomaxilar, cerca de la glabela, que no es prominente.
- El margen supraorbitario es delgado o mínimamente desarrollado.
- Hay un vaciamiento leve de la escama frontal, justo detrás de la glabela.
- La cara es mucho más larga que la de A. africanus.
- El perfil occipital es alto y aplanado.
- Hay una pequeña cresta sagital posterior en los machos.
Enlaces:
Metopic suture of Taung (Australopithecus africanus) and its implications for hominin brain evolution. Dean Falk et al (2012).Gómez Castanedo, A: (2012), "Los orígenes del género Homo. Una exploración de la innovación como motor evolutivo". Revista Nivel Cero, 13. Santander.
Bibiliografía:
- Dart, Raymond A., and A. Salmons. "Australopithecus africanus: The Man-Ape of South Africa." (1925).
- Dart, Raymond A. "The Makapansgat proto‐human australo‐pithecus prometheus." American journal of physical anthropology 6.3 (1948): 259-284.
- Dart, Raymond A. "Innominate fragments of Australopithecus prometheus."American journal of physical anthropology 7.3 (1949): 301-334.
- Broom, Robert. "Another new type of fossil ape-man." Nature 163 (1949): 57.
- Broom, Robert, and J. T. Robinson. "Further evidence of the structure of the Sterkfontein ape-man Plesianthropus." Transvaal Museum, Pretoria. Part I(1950).
- Tobias, Phillip V. "Recent Studies on Sterkfontein and Makapansgat and Their Bearing on Hominid Phylogeny in Africa." Goodwin Series 2 (1974): 5-11.
- Laitman, Jeffrey T. "Taung revisited: An examination of the past, present, and future of hominid evolution." Current anthropology 27.1 (1986): 78-80.
- Rayner, R. J., B. P. Moon, and J. C. Masters. "The Makapansgat australopithecine environment." Journal of Human Evolution 24.3 (1993): 219-231.
- Clarke, Ronald J., and Phillip V. Tobias. "Sterkfontein Member 2 foot bones of the oldest South African hominid." Science 269.5223 (1995): 521-524.
- Sponheimer, Matt, et al. "Hominins, sedges, and termites: new carbon isotope data from the Sterkfontein valley and Kruger National Park." Journal of Human Evolution 48.3 (2005): 301-312.
- Herries, Andy IR, et al. "A multi-disciplinary perspective on the age of Australopithecus in Southern Africa." The paleobiology of Australopithecus. Springer Netherlands, 2013. 21-40.
- Clarke, Ronald. "Australopithecus from Sterkfontein Caves, South Africa." The paleobiology of Australopithecus. Springer Netherlands, 2013. 105-123.
Te puede interesar:
|
Dating for everyone is here: ❤❤❤ Link 1 ❤❤❤
ResponderEliminarDirect sexchat: ❤❤❤ Link 2 ❤❤❤
ur. .
Esperame
Eliminar