 |
| Cladograma más parsimonioso para heidelbergensis, neanderthalensis y sapiens. Mounier y Caparros, 2015. |
- Heidelbergensis.
- Rodhesiensis.
- Erectus petraloniensis
- Archanthropus europeaus petraloniensis
- Swanscombensis
- Soloensis
- Etc.
 |
| Extensión geográfica del heidelbergensis. Manzi, 2016. |
En 1960, Clark Howell llegó a la conclusión de que los homínidos europeos eran muy particulares y diferentes a los del norte de África y, por supuesto, muy distintos a Homo erectus. Sin embargo, otros investigadores optaron por incluir a los fósiles europeos en erectus, aún reconociendo que se alejaban de la morfología clásica de esta especie.
 |
| Mauer 1. Schoetensack, 1908. |
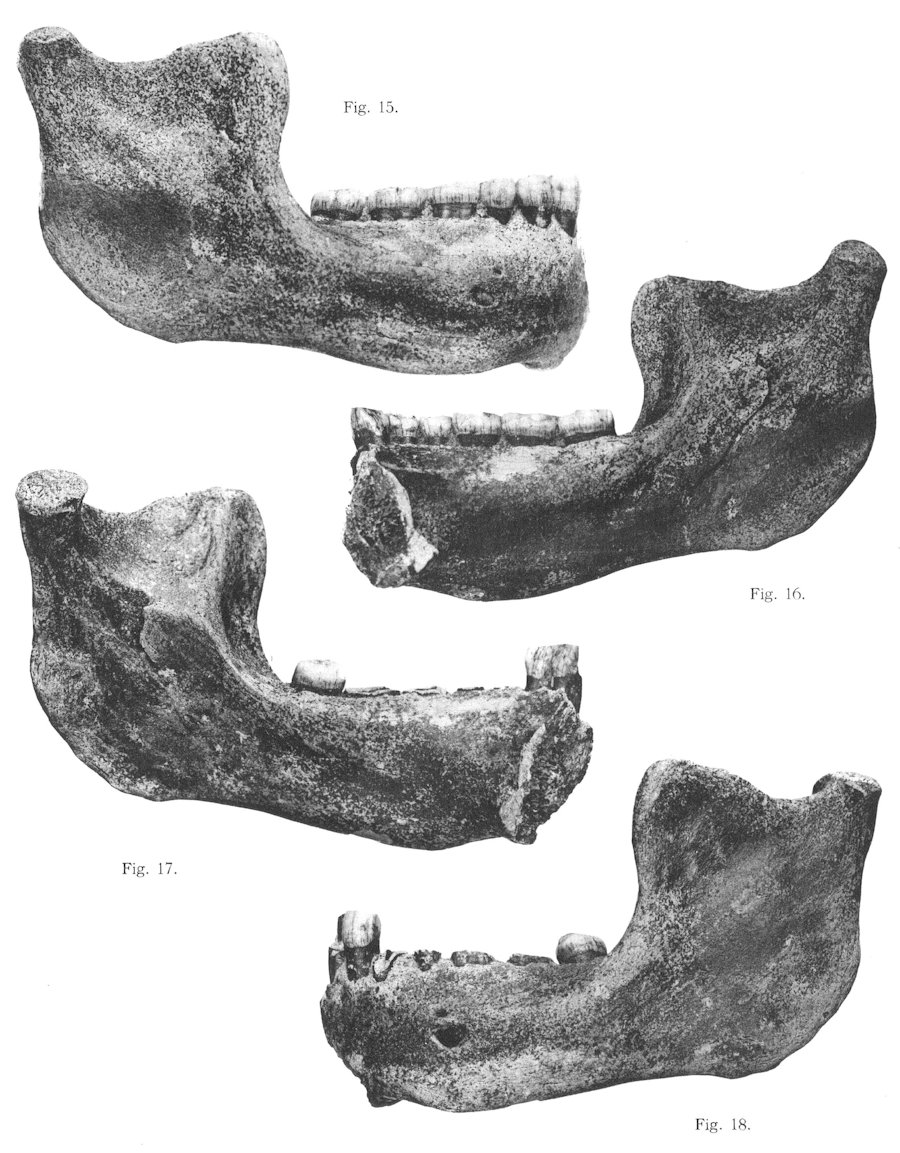
Chris Stringer, Günter Brauer y Phillip Righmire propusieron en los años 1980 que nuestra especie y los neandertales tuvimos un antepasado común, cuya cronología estimaron de manera provisional en medio millón de años. Homo erectus no parecía el mejor candidato y la cronología coincidía más o menos con la de la mandíbula de Mauer, holotipo de Homo heidelbergensis, Otto Schoetensack, 1908. En esta especie se incluyeron algunos fósiles africanos, todos los fósiles europeos y varios de China entre 600-200 ka, entre ellos Arago, Petralona, Broken Hill, Yunxian y Bodo. Algunos investigadores consideraron que los fósiles africanos formaban una especie separada (Homo rhodesiensis). La muestra más amplia disponible corresponde a la Sima de los Huesos de Atapuerca, más cercana a los neandertales que al ejemplar de Mauer.
Uno de los mayores problemas consiste en que disponemos de muy pocas mandíbulas para compararlas con la de Mauer y además para algunos investigadores, como Yoel Rak, el espécimen de Mauer es único. Tattersall, por el contrario, encuentra afinidades con las mandíbulas de Arago, sobre todo en los dientes. Juan Luis Arsuaga ha propuesto la designación de un nuevo holotipo, que conserve el cráneo completo. Berhane Asfaw ha comunicado el hallazgo, todavía no publicado, de fragmentos de cráneo y mandíbula de 32 individuos en la región de Middle Awash, Etiopía, datados en 300 ka que recuerdan los hallazgos de Arago y de Herto. En un análisis cladístico, Mounier y Caparros (2015) consideran que la especie Homo heidelbergensis no es coherente, no está bien apoyada y es equívoca.
Yacimientos europeos con restos humanos datados entre ca 800-350 ka y presencia de contextos achelenses. Peretto et al, 2015.
1. Gran Dolina TD6
2. Mauer
3. Boxgrove
4. Isernia La Pineta
5. Caune de l’Arago
6. Sima de los Huesos
7. Ceprano
8. Fontana Ranuccio
9. Terra Amata
10. Visogliano
11. Venosa–Notarchirico
12. Pofi
13. Bilzingsleben
14. Vértesszõlõs
|
- Atribuirlos al grado erectus.
- La solución parsimoniosa es colocarlos todos dentro de erectus.
- David Lordkipanidze et al (2013), Adam van Arsdale y Milford H. Wolpoff (2012) y otros investigadores como Finlayson son partidarios de un único linaje humano muy variable. Las diferencias fenotípicas serían adaptaciones geográficas y no evidencias de diversidad de especies.
- Las evidencias genéticas indican hibridación entre los diferentes grupos humanos. Según la definición de especie biológica de Ernst Mayer, esto implicaría la pertenencia a la misma especie.
- Para Stringer (1993) la falta de rasgos derivados propios del erectus en los ejemplares de Atapuerca, confirma su tesis de que nunca hubo erectus en Europa y sugiere que pertenecen ya al taxón de los neandertales.
- Atribuirlos a la especie Homo heidelbergensis. En 1983 Stringer, en un estudio del cráneo Petralona propuso H. heidelbergensis como la especie madre afro-europea de H. neanderthalensis y H. sapiens. Este punto de vista fue posteriormente apoyado y desarrollado por Rightmire (1997) para quien heidelbergensis es un taxón válido que quizá sea el último antecesor común de neandertales y humanos modernos, con una evolución por separado en Europa y África. Para Ward y Stringer la población europea lleva hasta los neandertales y la africana a los sapiens.
- Atribuir los restos africanos a rhodesiensis (luego a helmei) y los europeos a:
- heidelbergensis. Creada en 1907 por Otto Schoetensack. No todos los fósiles de heidelbergensis presentan rasgos neandertales en la misma parte anatómica El equipo de Atapuerca (2011) considera que heidelbergenesis es un grado, un conjunto de grupos con evolución en paralelo. Los neandertales pudieron surgir del grupo de Atapuerca o de otro.
- neanderthalensis. Stringer (1993) y Arsuaga consideran que la gran variación de la muestra de Atapuerca hace difícil seguir sosteniendo el taxón heidelbergensis porque esta especie y los neandertales se solaparían. Como mucho heidelbergensis sería un ancestro exclusivo de los neandertales. Los homínidos europeos del Pleistoceno Medio formarían una cronoespecie y serían antecesores solo de los neandertales, que derivarían por anagénesis. Esto obligaría a extender neanderthalensis a través de un periodo muy largo, en el que hubo cambios morfológicos notables.
- Según el Modelo de Acumulación (Accretion Model) propuesto por Hublin, podemos distinguir varias etapas o saltos (Dean et al, 1998):
- Preneandertales tempranos: Mauer, Petralona, Arago.
- Preneandertales: Reilingen, Steinheim, Swanscombe, Sima del Huesos de Atapuerca. Este grupo muestra el comienzo de los superciliares de doble arco y prognatismo mediofacial.
- Según J. L. Arsuaga et al (2014), la población de la Sima de los Huesos no puede clasificarse ni como heidelbergensis ni como neanderthalensis.
- El análisis génetico realizado por Meyer et al (2015), confirmó la pertenencia de la población de la Sima de los Huesos al linaje neandertal.
- Neandertales tempranos: Biache, Ehringsdorf, Saccopastore, Kaprina y algunos de Shanidar. Aparición del moño occipital.
- Neandertales clásicos: Feldhofer 1, Gibraltar 1 (Forbes Quarry), La Chapelle-aux-Saints, La Ferrassie, Amud y el resto de los ejemplares de Shanidar. Incremento del prognagtismo mediofacial y sulcus orbital profundizado.
- Para Aguirre y Lumley (1977), en el Pleistoceno Medio existieron tres grupos genética y geográficamente separados, que no pueden ser considerados erectus (que quedaría restringido a Java y no contribuiría a la aparición de neandertales ni sapiens), aunque su cercanía supone un argumento a favor de la hipótesis multirregional erectus-sapiens:
- Lejano Este (Zhoukoudian). La posibilidad de coexistencia en Java de erectus y sapiens ha sido propuesta por Swisher y colaboradores (1996). Para ellos, los erectus sobrevivieron en Java al menos 0,25 mda después de haberse extinguido en China. Sugieren incluso que erectus y sapiens pudieran intercambiar genes. De clasificar el espécimen hallado en Jinniu Shan (0,2 mda) como sapiens, las dos especies coexistirían también en China.
- Magreb.
- Grupo anteneandertaliense: Mauer, Arago, Atapuerca, Montmaurin, Bañolas.
- Howell (1999) distinguió las paleodemos:
- Mauer/Arago
- Petralona/Atapuerca Sima
- Tattersall (2011), con los fósiles europeos, hace los siguientes grupos:
- Relacionados con Homo erectus: Swanscombe, Verteszöllos, Bilzingsleben y Biache-Saint-Vaast.
- Homo heidelbergensis: Mauer, Arago, Petralona, Ceprano y Boxgrove. A heidelbergensis pertenecerían también los hallazgos de Kabwe y Bodo en África y Dali y Jinniushan en China.
- Un último grupo de filogenia desconocida: Steinheim, Reilingen y Sima de los Huesos.
- Adrián Pablos et al (2013) han comparado 25 astrágalos de 14 individuos (once adultos y tres inmaduros) de la Sima de los Huesos (Atapuerca), con los correspondientes a sapiens y neanderthalensis. Algunos caracteres comunes con los neandertales son consistentes con la hipótesis de que la población de la Sima de los Huesos y los neandertales son grupos hermanos.
- A partir de OIS 6 (175 ka), todos los hallazgos europeos corresponden a neanderthalensis.
- Vandermeersch y M. D. Garralda (2011) distinguen en Europa dos linajes:
- erectus tardío.
- Preneandertales u Homo heidelbergensis. Este taxón y neanderthalensis son cronoespecies.
- Reconocen la dificultad de diagnosticar Homo heidelbergensis.
- No puede ser excluido un intercambio genético entre estos grupos. Un escenario probable incluiría además múltiples migraciones de pequeños grupos desde varios puntos.
- Mounier y Caparros (2015).
- Una serie de especímenes del Pleistoceno Medio presentan una posición filogenética incierta, por su heterogeneidad: Arago, Ceprano, Petralona, Broken Hill 1, Bodo y Dali. Forman una politomía que parte del último ancestro común entre los humanos modernos y los neandertales.
- Sima de los Huesos y Steinheim forman parte del clado neandertal.
- Bermúdez de Castro et al (2016). Bermúdez de Castro et al (2018).
- Los especímenes de Mauer, Arago, Sima de los Huesos, Steinheim, Swanscombe o Pontnewydd pueden proceder de diferentes migraciones desde el suroeste de Asia, a partir de una población madre común que evoluciona en esta última región.
- Los primeros colonos pueden identificarse por su morfología más plesiomórfica, mientras que los colonos más recientes se parecen más a los neandertales.
- La variabilidad observada en los especímenes del Pleistoceno Medio europeo podría explicarse por una historia compleja de aniquilamiento y recolonización y la acción de diversos procesos genéticos, incluidos la deriva, el efecto fundador, la hibridación en distintos grados y la adaptación direccional (Vialet et al, 2018).
- La especie H. heidelbergesis debe ser eliminada de la filogenia. Los especímenes europeos del Pleistoceno Medio podrían ser tentativamente incluidos en dos o más ramas del clado neandertal.
- Mercedes Conde-Valverde et al (2018)
- A partir de los fósiles de Aroeira y Sima de los Huesos, predicen la existencia de un paleodeme en la Península Ibérica durante MIS 13, caracterizado por la presencia al menos de una región glabelar prominente, un proceso postglenoideo bien desarrollado y la forma del giro basal coclear.
 |
| Esquema de la evolución humana en Europa. H. antecessor podría representar una rama europea cercana al último antecesor común (LCA) de neandertales y sapiens. La especie H. heidelbergesis debería ser retirada de la filogencia humana y los especímenes del Pleistoceno Medio podrían inclurise en dos o más ramas del clado neandertal (1, 2, y 3 en la figura). Bermúdez de Castro et al, 2016. |
Según María Martinón et al (2010) el aislamiento geográfico se produjo
hace 800 ka debido al desierto Sahara-Arábico.
La migración a Europa traería consigo el Modo 2. Gracias a esta
tecnología y a la caza organizada de grandes herbívoros (Will Roebroeks, 2001),
heidelbergensis pudo ocupar zonas
septentrionales de Europa hasta enconces escasamente pobladas. La densidad de
población aumentó drásticamente en toda Europa.
Evolución de Homo durante el interglaciar Mindel-Riss
 |
Árboles evolutivos alternativos que muestran las relaciones entre Homo erectus y los homínidos del Paleolítico Medio, los neandertales y los HAM.
a. Las poblaciones del Pleistoceno Medio y posteriores se agrupan en un grado sapiens.
b. Existen al menos dos linajes además de erectus y HAM. Una rama europea, comparte apomorfias con los neandertales y puede distinguirse de un segundo linaje localizado en África (Homo rhodesiensis).
c. Las diferencias morfológicas entre los especímenes europeos y africanos son pequeñas y pueden atribuirse a variación geográfica e intraespecífica. Si la mandíbula de Mauer es el hipodigma, el nombre apropiado para esta especie, es Homo heidelbergensis
|
Modelo de Fuentes y Sumideros
El conjunto de la Sima de los Huesos presenta una morfología derivada neandertal más pronunciada que los ejemplares de Mauer, Arago o Ceprano. Para explicar esta variabilidad, Dennell, Martinón-Torres y Bermúdez de Castro (2011), Bermúdez de Castro y Martinón-Torres (2013), propusieron un modelo de población que se basa en fuentes y sumideros demográficos. Un pequeño número de fuentes en el sur del continente repueblan áreas más al norte durante los interglaciares, dando lugar a grupos sumidero. Estos grupos sumidero se extinguirían durante las épocas glaciares. De esta forma la variabilidad vendría favorecida por sucesivas dispersiones, fragmentaciones y recombinaciones de las poblaciones.
Diferentes vistas de algunos de los cráneos más significativos. Francesco Mallegni (2011).
 |
| Cráneos correspondientes al Pleistoceno Temprano y Medio |



No hay comentarios:
Publicar un comentario