 |
| LB 1. Homo floresiensis |
En la Isla de Flores, el yacimiento lítico de Mata Menge ha sido datado en más de 1 Ma (
Brumm et al, 2010). La caída de más de 150 m de las líneas de costa, pudieron haber favorecido istmos estrechos y corredores para la migración. Según
Robin W. Dennell et al (2013) la fauna de mamíferos, aves y reptiles de Flores procedía de varios lugares, incluyendo Java, Sulawesi y Sahul. Muchos de los taxones terrestres eran capaces de flotar o nadar, mientras que, si consideramos las corrientes predominantes, los roedores y los
Homo probablemente navegaron accidentalmente desde Sulawesi.
- Gerrit D. van den Bergh et al (2016) han comunicado el hallazgo de 41 artefactos líticos in situ asociados con restos fósiles de megafauna (Bubalus sp., Stegodon y Celebochoerus) en la trencha 4 (T4) y otros cuatro de pequeño tamaño en la sub-unidad E2, más antigua, en Talepu, Cuenca Walanae, noreste de Maros, isla de Sulawesi. Los artefactos no han podido ser clasificados en ninguna industria en particular. La edad mínima es de hace 118 ka mientras que los artefactos de E2 pueden llegar hasta hace 194 ka.
 |
| Posibles rutas hasta Flores de los antepasados del Homo floresiensis |
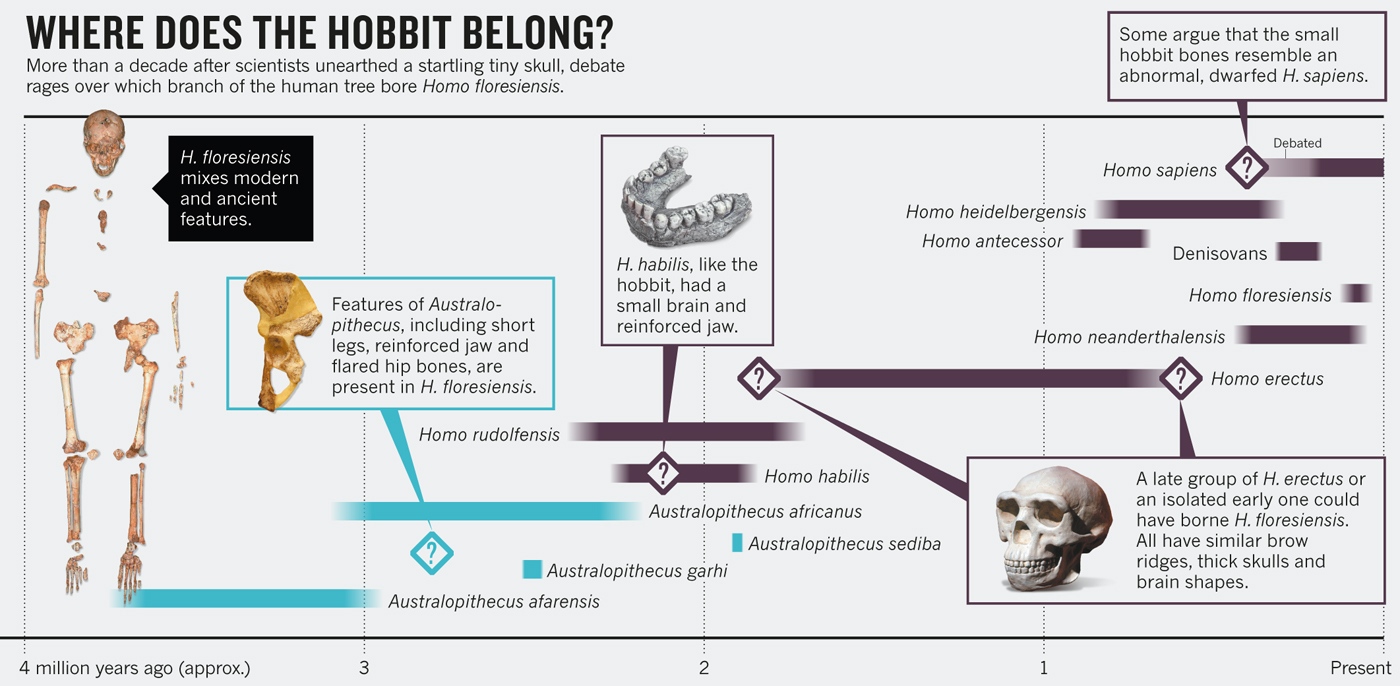
El 28 de octubre de 2004 Thomas Sutikna y Mike Morwood anunciaron el hallazgo de los restos de un homíndo de pequeño tamaño (LB1) en la cueva de Liang Bua, Isla de Flores, asociados a instrumental lítico sofisticado.
La descripción corrió a cargo de Peter Brown quien atribuyó los restos a una nueva especie: Homo floresiensis. Vulgarmente, se le conoce como
Hobbit (del
Señor de los Anillos).
Este descubrimiento planteó una paradoja: Con un cerebro de tamaño parecido al de un chimpancé, la nueva especie mostraba indicios de inteligencia avanzada como por ejemplo la caza con armas de piedra, despedazar cadáveres o controlar el fuego.
- La hipótesis del sesgo de cautividad relaciona la frecuencia de uso y variedad de las herramientas con el tamaño y la interconexión de las poblaciones y no con la capacidad cognitiva (Michael Haslam, 2013).
En octubre de 2005 Morwood et al., hallaron nuevos restos (entre ellos, algunos huesos de la mano omóplato y mandíbula del mismo individuo, LB6).
Fueron datados entre 95-12 ka. Los nuevos restos descartaban que el tamaño de floresiensis se debiese a una patología; además, sus caracterísiticas estaban alejadas de la línea del erectus y el sapiens, Morwood y Debbie Argue propusieron que floresiensis podría descencer de una especie más antigua, un australopitecino o quizá habilis, que migró de África antes que los ancestros del erectus hiciesen lo mismo (2,5 Ma).
 |
| Hallazgos de Homo floresiensis |
Floresiensis se define por:
- Estatura sobre 1,06 m. 32,5 kg.
- Muy pequeña capacidad craneal (426 cc; Daisuke Kubo et al, 2013). Bóveda craneana más gruesa que en Australopitecus y similar a H. erectus y H. sapiens. La bóveda craneana es baja y angosta en comparación con la del hombre actual y aun en comparación con la de erectus. La flexión basicraneal que es un rasgo determinante en la hominización, es de 130°, intermedia entre erectus (141°) y el hombre actual (127°). Su estatura total y su volumen craneano ofrecen valores que están por fuera del rango de variación de los homínidos modernos. Coincidentemente, el grado de encefalización –expresado como cociente entre el volumen endocraneano y la longitud femoral (gran indicadora, esta última, de la talla total)– arroja para floresiensis un valor (5,9) menor al de erectus (6,9) y aún menor al de afarensis (6,1), mientras que en los homínidos actuales dicho valor gira en torno a 7,5. Para Dean Falk, los rasgos cerebrales preservados en su cráneo sugieren que el homínido de Flores podría ser capaz de realizar tareas cognitivas avanzadas.
- El cerebro del Homo floriensis posee lóbulos temporales bastante amplios, regiones del cerebro asociadas con la comprensión del habla y el oído. Aún más sorprendente es el hecho de que posee lóbulos frontales del cerebro que tienen circunvoluciones con muchos pliegues, áreas del cerebro directamente debajo de la frente que están implicados en un reconocimiento superior. La región más circunvolucionada es la parte que se proyecta más allá del lóbulo frontal, llamada el polo frontal. Falk identifica esta región como área 10 de Brodmann, la cual está expandida en humanos modernos y está relacionada con la toma de iniciativas e ideación de planes de acciones futuras que son componentes clave de la cognición elevada. Esta área alargada sugiere que Homo floresiensis pudo confeccionar las herramientas de piedra que se encontraron en las proximidades, las cuales son más típicas de humanos prehistóricos modernos que de antiguos homínidos como erectus.
- Arcos superciliares de tamaño moderado.
- Mandíbulas salientes. La sínfisis mandibular no se resuelve en un verdadero mentón como en sapiens, sino que presenta inclinación en sentido póstero-inferior del eje sinfisario, como en los australopitecinos o el habilis. Protuberancias o contrafuertes mandibulares, comunes en los homínidos tempranos pero no en el HAM. Hueco entre el extremo de la fila de dientes y la sección posterior de la mandíbula.
- Sin las especializaciones masticatorias ya presentes en Australopithecus y Paranthropus. Muelas relativamente grandes. Raíces dentales diferentes al HAM.
Según
David J. Daegling et al (2014) la mandíbula de
floresiensis es relativamente fuerte (comparable a la de los australopitecinos) en términos de rigidez en la torsión y la flexión transversal, pero es relativamente débil bajo flexión parasagital. Las cargas masticatorias se redujeron en relación con los australopitecinos, pero se mantuvieron relativamente elevadas con respecto a los HAM.
- Yousuke Kaifu et al (2015) han analizado los restos dentales de Homo floresiensis.
- Conjunto C/P3 y otras características primitivas, similares a Homo erectus.
- C1: hombro distal bajo.
- P3: cresta transversal. Corona larga y asimétrica en dirección MD. Esmalte bucal hinchado (cíngulo). Raíz bifurcada.
- P4: cresta transversal. Cara lingual de la corona, bien desarrollada.
- M1: cresta media del trigónido.
- Tamaños equivalentes de M1 y M2.
- H. floresiensis
- Morfología molar avanzada, incluso en comparación con los HAM.
- Apomorfias de H. floresiensis:
- Gran tamaño relativo y morfología oclusal única del P3.
- Primeros molares extremadamente pequeños. No está clara la razón por la que H. floresiensis experimentó una evolución tan singular.
- Esta combinación de rasgos es desconocida hasta el momento en otras especies de Homo. La dentición de H. floresiensis no es una versión a escala reducida de Homo anteriores.
- Prognatismo facial. Frente huidiza.
- Tiene arcos supraorbitarios marcados, aunque no llega a formar el clásico torus.
- Meatus auditivo externo pequeño y redondeado, algo alargado verticalmente, con margen timpánico muy fino, que recuerda a los australopitecinos.
- Bajos ángulos glenoideo y axilo espinal del omóplato. Recuerda a Turkana boy (ergaster) y australopitecinos.
- Bajo ángulo de torsión del húmero.
- Clavícula corta.
- Muñeca primitiva, que recuerda la de los australopitecinos o la de habilis.
- Trapezoide acunado.
- El lado cubitar de la articulación del segundo metacarpo, orientado sagitalmente
- La articulación del escafoide, algo triangular
- Falta el área de expansión palmar carcterística del último Homo y del Pleistoceno Medio, que permite la pinza de precisión.
- Escafoide y capitate plesiomórficos.
- Piernas cortas, como las de los australopitecinos.
- Pie excepcionalmente largo en relación al fémur y a la tibia. Hállux completamente aducido. Otras características sin embargo, recuerdan a los simios. Junto con otras características, los rasgos del pie indican una forma de desplazamiento biomecánicamente distinta a la del sapiens (W. L. Jungers et al, 2009).
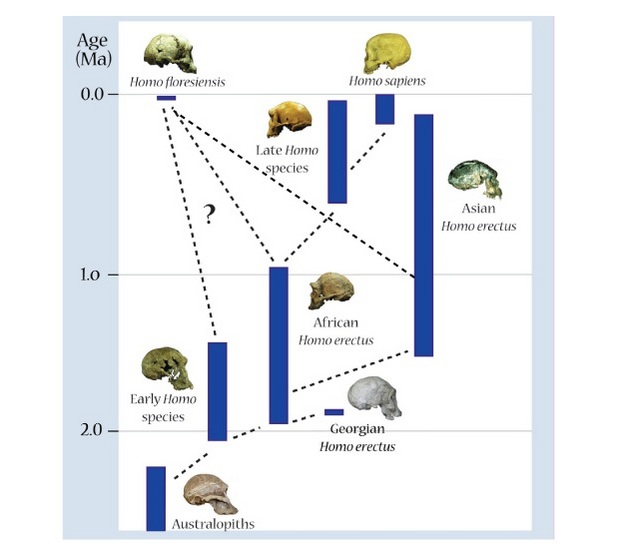 |
| Posibles escenarios evolutivos para Homo floresiensis |
Teorías sobre el origen del Homo floresiensis
Según el
análisis cladístico efectuado por D. Argue et al (2012), los resultados producen dos árboles filogenéticos igualmente parsimoniosos. El primero sugiere que
H. floresiensis es un homínido temprano que surgió después de
Homo rudolfensis, pero antes de que
H. habilis. El segundo árbol indica que
H. floresiensis surgió después de
Homo habilis.
- Homo floresiensis no es humano.
- Los restos hallados corresponden a casos patológicos. Estas propuestas no explican como se pudo mantener la patología durante tantos miles de años y la ausencia de espécimenes sin la referida patología.
- Oxnard et al (2013) han criticado los estudios y conclusiones de Brown et al, quienes consideran que floresiensis fue un descendiente de erectus, en una línea evolutiva afectada por el enanismo insular
- Para Teuku Jacob y Robert Martin, LB1 se trataba de un sapiens que padeció microcefalia. Oxnard et al, 2010, Brown 2012 los consideran casos de cretinismo.
- Opinan lo mismo Obendorf et al (2008) en base al tamaño de la silla turca y Vannucci el al (2011) en base a un análisis de tomografía computerizada.
- Para Falk et al. 2009, (ver Hawks 2007), estamos ante un caso de síndrome de Laron.
- Robert B. Eckhardt et al (2014), Robert B. Eckhardt et al (2015), consideran que las principales características (volumen cerebral y estatura) de LB1 están subestimadas ya que se ha pasado por alto la asimetría y la desproporción del ejemplar, que son signos de desarrollo anormal. El ADN extraído de los huesos de LB1 es demasiado fragmentario para permitir un diagnóstico, pero los fragmentos recuperados hasta el momento son de tipo humano moderno. Según los autores, incluso en ausencia de un diagnóstico sindrómico en particular, las características que definen a LB1 no cumplen con los criterios formales para un espécimen tipo de una nueva especie.
- Para Maciej Henneberg et al (2014), la asimetría facial, el pequeño volumen endocraneal, la braquicefalia, los fémures desproporcionadamente cortos, los pies planos, y otras numerosas características de LB1 son altamente diagnósticas de síndrome de Down, uno de los trastornos del desarrollo que aparecen más frecuentemente en los seres humanos y también se documenta en los hominoideos como los chimpancés y los orangutanes. Consideran que el mentón, cuya presencia es un importante caracterísitica definitoria del HAM, está a menudo ausente entre los australo-melanesios.
- Michael Carrington Westaway et al (2015) advierten que los estudios sobre la ausencia de mentón de los autralo-melanesios no han sido revisados o han sido criticados severamente y consideran, de acuerdo con Brown y Tomoko, que la ausencia de barbilla en LB1 y LB6 imposibilita su atribución a H. sapiens. Otras características de la barbilla confirman esta imposibilidad.
- H. floresiensis experimentó una dramática reducción de tamaño del cerebro y del cuerpo a partir de la condición de grado erectus en un entorno insular aislado (enanismo insular).
- Una primera hipótesis explicativa recurrió al enanismo insular (dwarfism), fenómeno evolutivo observado en las islas en muchas ocasiones consistente en una gran reducción de tamaño, como forma de sobrevivir en un entorno pobre. Para Peter Brown y colegas, floresiensis fue un descendiente de erectus, en una línea evolutiva afectada por el enanismo insular.
- Hay detalles de la antomía de LB1 que recuerdan al erectus: El neurocráneo, la frente plana e inclinada, el espesor craneal de los huesos, la cara corta y plana, el toro occipital, la fisura del mastoidos y el hombro, pero tanto el pequeño tamaño del cuerpo y el tamaño del cerebro se encuentran fuera del rango esperado para la especie (Brown et al. , 2004; Falk et al. , 2005; Baab y McNulty, 2009; Baab, McNulty y Harvati, 2013). Igualmente, la dentición es similar a la de erectus (Kaifu et al, 2015).
- Según esta teoría, los ancestros de los Homo floresiensis de alguna manera navegaron hasta Flores, donde se produjo una reducción en el tamaño del cuerpo (Meijer et al, 2010).
- H. floresiensis deriva directamente de una forma más primitiva de encéfalo pequeño como el Homo habilis (aprox. 600 cc) o incluso Australopithecus (aprox. 400 cc).
- No existe ninguna evidencia fósil en Asia de estas especies.
- Las evidencias de la mandíbula, los dientes y el esqueleto postcraneal apoyan esta hipótesis (Argue et al, 2009). El tamaño y la morfología de los dientes y la mandíbula comparten más semejanzas con Australopithecus y los primeros Homo que con los erectus (Brown y Maeda, 2009). En particular, las piernas muy cortas (con respecto tanto a los brazos como a los pies) son un patrón común en los monos y los australopitecinos pero no en erectus. LB1 también era desproporcionadamente pesado para su altura, como Lucy (Australopithecus afarensis) (Jungers y Baab, 2009). Los huesos del carpo (muñeca) se parecen más a los de los chimpancés a los de los humanos modernos (Tocheri et al, 2007). La morfología del pie, también conserva varias características muy primitivas: una larga parte delantera del pie con los dedos curvados y la falta de un arco longitudinal medial (Jungers et al, 2009).
- H . floresiensis no sigue la tendencia alométrica del Homo del Pleistoceno Medio, los neandertales y el HAM (Juan Antonio Pérez-Claros, Juan Manuel Jiménez-Arenas y Paul Palmqvist, 2015)
- Si H. floresiensis derivó del HAM en condiciones insulares, tuvo que ser a través de transposiciones laterales. Esto significa que debe ser pedomórfico en tamaño y peramórfico en forma con respecto al HAM.
- Sin embargo, es más fácil conectar LB1 con los habilinos por escalamiento ontogénico, lo que significa que LB1 sería pedomórfico tanto en tamaño como en forma. Esta es una explicación más parsimoniosa.
 |
| Teorías sobre el origen de Homo floresiensis |
Karen L. Baab ha revisado la morfología de
Homo floresiensis
- La morfología del cráneo y neurocráneo de LB1 es más parecida a H. erectus que a H. habilis.
- En particular, se parece al subadulto de Dmanisi D2700, en parte debido a los patrones de escala.
- El grosor de los huesos craneales recuerda también a H. erectus, sin embargo, este apoyo es débil debido al pequeño tamaño de la muestra correspondiente a los primeros Homo.
- En la comparación de 67 rasgos craneofaciales, Kaifu et al (2011) interpretaron que el patrón de floresiensis es consistente con una ascendencia de los erectus tempranos de Indonesia.
- La forma mandibular general ha sido descrita como similar a la de los primeros erectus africanos y a la población de Dmanisi, aunque con algunas características más primitivas.
- La escala M1> M2> M3 es compartida con erectus y sapiens peron no con los primeros Homo.
- La sínfisis mandibular fuertemente inclinada es compartida con algunos ejemplares de la población de Dmanisi, erectus y australopitecinos gráciles, pero no con los primeros Homo ni con los erectus de Indonesia.
- Las coronas molares mandibulares son más pequeñas que las de los primeros Homo y australopitecinos.
- La prominencia alveolar lingual bien desarrollada es compartida únicamente con los erectus africanos.
- La orientación de la cresta transversal distolingual en el P3 es una característica derivada compartida con Dmanisi y los erectus africanos.
- La falta de elementos postcraneanos de los primeros Homo y de los erectus, especialmente de los asiáticos, hace difícil la interpretación de rasgos compartidos con floresiensis.
- La prominente tuberosidad navicular de LB1 es más primitiva que la observada en OH8 y parece indicar que carecía de arco longitudinal medial, aunque también Jinniushan y los neandertales muestran una tuberosidad navicular prominente, por lo que la interpretación es controvertida.
- El bajo grado de torsión de la cabeza del astrágalo en LB1 está más cerca de los monos africanos que de los humanos, aunque se han reportado valores similares para OH8.
- En KNM-WT 15000 y LB6, la fosa glenoidea se orienta lateralmente en lugar de cranealmente, lo que constituye una morfología derivada desconocida en los primeros Homo. En Dmanisi en cambio, se orienta más cranealmente.
- Muchos rasgos anatómicos de floresiensis son compartidos tanto con erectus como con los primeros Homo.
- Kaifu et al (2011) identificaron al menos 20 rasgos craneales de LB1 que también se encuentran en habilis y algunas poblaciones de erectus, incluyendo un débil o nulo desarrollo de las quillas coronal y sagital, un escama temporal grande con una sutura parietomastoidea corta, la presencia de un proceso postglenoideo, una fosa mandibular poco profunda y un surco supraorbital que no se distingue con claridad.
- La corona asimétrica y alargada mesiodistalmente del P3 inferior, documentada en LB1 y LB6 es común en los australopitecino y en Homo temprano y, aunque es rara en erectus, se observa en Dmanisi. La raíz bifurcada es común en los australopitecinos y se observa también en algunos erectus tempranos.
- LB1 presenta un P4 inferior con la raíz dividida. Este tipo de raíz (raíz de Tomes), con la cámara pulpar simple en forma de cinta arqueada 180º o más (C-shaped) aparece por vez primera en habilis coincidiendo con la reducción en tamaño de los dientes, pero mientras que para los P3 inferiores se han documentado en los primeros Homo, en los P4 inferiores son poco comunes en el registro fósil, pero están presentes en los primeros Homo y en los erectus de África e Indonesia.
- Los huesos del carpo de floresiensis muestran muchas características observadas en los australopitecinos y en los monos, pero no en los neandertales y en los HAM, como un trapecio en forma de cuña, y un capitate con un cuello entallado y una faceta del escafoides que se extiende sobre el cuello en forma de J. La morfología carpal es desconocida para los primeros Homo y erectus.
- LB1 / LB6 comparten baja torsión humeral y bajo ratio claviculohumeral con erectus y Hominini anteriores.
- La poca profundidad dorsal del surco troclear de LB1, es una condición derivada en la dirección del sapiens que aparece tanto en los Homo tempranos como en los erectus. Las alas abiertas hacia fuera en la faceta del maleolo externo parecen conservarse en la mayor parte de los fósiles antreriores al sapiens. El hallux es bastante corto en relación a los otros dedos, lo que constituye una característica primitiva, similar a la que aparece en Dmanisi. En LB1, Dmanisi y los Hominini de Illeret y está aducido al resto del dedos en contraste con los Hominini basales y Burtele o el grado intermedio documentado para los australopitecinos. Sin embargo, las huellas de Laetoli y OH8 también muestran un hallux aducido.
Este análisis revela que el antecesor de
floresiensis debió de ser
erectus, habilis u otra especie relacionada.
- Las características primitivas, las morfologías más similares a los australopitecinos que a Homo, han de corresponder a convergencias evolutivas. Tal vez la condición primitiva de la rama y la sínfisis mandibular se deba a un cambio hacia una masticación más poderosa. La morfología de la tuberosidad navicular, la ausencia de arco longitudinal y el pequeño tamaño del cerebro son otras reversiones tanto con respecto a erectus como a Homo temprano.
- Si floresiensis es descendiente de erectus, aumentó entonces la robustez relativa de las extremidades superiores, lo que permite especular con la presencia de comportamientos más arbóreos, si bien las características del pie no apoyan esta hipótesis. La masa y la estatura se redujeron sustancialmente (enanismo insular). Se ha especulado con una migración no documentada hasta el momento, de erectus tempranos, relacionados con Dmanisi, de tamaño relativamente pequeño, al sudeste de Asia, de la que provendría floresiensis.
- Si floresiensis es descendiente de habilis, muchas características del cráneo y algunos rasgos dentales tuvieron que evolucionar en paralelo en floresiensis y erectus (homoplasias). Algunos de estos rasgos pudieron evolucionar en forma correlacionada, lo que deja abierta la posibilidad de que los rasgos compartidos surgieran gracias a la selección de uno o unos pocos en presencia de presiones similares. Si OH 62 midió unos 118 cm, la estatura de LB1 de unos 106 cm requirió solo una pequeña reducción.
- Esta ascendencia supondría una migración Out of Africa no documentada hasta el momento.
La hipótesis de que
erectus es el ascendente de
floresiensis parece más parsimoniosa que una evolución en paralelo de los rasgos craneales y dentales compartidos por
erectus y
floresiensis.
Homo floresiensis

No hay comentarios:
Publicar un comentario