Se ha aceptado, en general, un modelo de rápida sustitución de los neandertales europeos autóctonos por los HAM inmigrantes, pero los últimos estudios nos proporcionan un panorama más complejo, caracterizado por un mosaico biológico y cultural que se prolongó durante varios miles de años en diferentes regiones.
- Según O. Jöris et al, 2011, en Europa, los fósiles neandertales con datación segura, son anteriores a 38 ka. Los humanos anatómicamente modernos con datación segura son posteriores a 35 ka.
- Según la datación de Higham et al (2006), los neandertales de Vindija tienen una antigüedad de 33-32 ka. Serían los últimos neandertales datados con seguridad.
- Trinkaus et al (2003, 2006) y Rougier et al (2007) dan una fecha de 40 ka para los primeros sapiens en el sureste de Europa. No llegaron al oeste de Europa hasta hace 37 ka (Dujardin, 2003; Henry Gambier et al, 2004; Wild et al, 2005). Alcanzaron el Atlántico hace 35 ka (Walker et al, 2008).
- N. J. Conard (2011), en el caso de Suabia, sugiere la llegada del sapiens hace 40ka por el corredor del Danubio. En ese momento, las poblaciones de neandertales eran extremadamente pequeñas.
- Ernsthofen, en la terraza baja del río Enns, alrededor de 300 m sobre el nivel del mar es el mayor yacimiento neandertal en la región del Danubio. Se han encontrado en superficie cientos de herramientas de alta calidad de cuarcita, cuarzo y otros materiales líticos en la típica técnica Levallois. Exactamente a la misma altura en las orillas del Danubio en Weinzierl, cerca de la ciudad de Perg, se halló un campamento de HAM con aproximadamente la misma datación. Los artefactos de los humanos modernos eran cuchillas y escamas de sílex y radiolaritas obtenidos de los depósitos de la terraza fluvial fabricadas mediante el método volumétrico. Aparecen también conchas de caracol perforadas y dientes perforados que probablemente fueron cosidos a la ropa o colgados de tiras de cuero.
- En la conferencia Biology of Genomes 2015, Fu el al presentaron el análisis genético de Oase 1, que ha resultado contener un 5-11% de genoma de origen neandertal, lo cual indica un antepasado neandertal entre 4 y 6 generaciones antes, en fechas muy cercanas a hace 40 ka (ver comentarios de Ann Gibbons).
- Según David Santamaría Álvarez y Marco de la Rasilla Vives (2013), en la Península Ibérica nunca hubo contacto entre neandertales y modernos ya que aquellos ya se habían extinguido cuando estos llegaron. C. Mallol et al (2012) son de la misma opinión.
- M. Cortés Sánchez et al (2011) aceptan una datación de 28 ka como límite de la supervivencia de poblaciones neandertales en el sur de la Península Ibérica (Cueva Gorham, Zafarraya...).
- Rachel E. Wood et al (2013) han publicado una nueva datación por el método del radiocarbono con protocolo de ultrafiltración, de los hallazgos de Cueva del Boquete de Zafarraya (Málaga) y Jarama VI (Sistema Central), dos de los últimos refugios neandertales. La nueva datación, de 45-50ka, de diferentes ejemplares, descarta la coincidencia con Homo sapiens en estos lugares. Igualmente, Bertila Galván et al (2014) en El Salt, con un límite de 45 ka.
- Jesús F. Jordá Pardo et al (2013) han analizado y datado (con ultrafiltración) los depósitos arqueológicos de La Güelga (Asturias, España), Según sus conclusiones, neandertales y sapiens coexistieron en aquellos lugares hace 40 ka. Los investigadores han detectado niveles con materiales atribuidos al hombre anatómicamente moderno (con restos tecnológicos del Auriñaciense) entre estratos con materiales producidos por neandertales (con instrumentos del Musteriense y del Chatelperronense). Es decir, los humanos modernos ocuparon durante un tiempo la misma cavidad que, antes y después, fue habitada por grupos de neandertales.
- Según Marín Arroyo et al (2018), la superposición en Cantabria entre el Chatelperroniense y el Auriñaciense no alcanza los mil años, de acuerdo con las dataciones obtenidas en 13 yacimientos.
- Tom Higham et al (2014) han construido cronologías robustas de 40 yacimientos musterienses y neandertales llegando a las siguientes conclusiones:
- El Musteriense finalizó hace 41,03-39,26 ka cal BP (al 95,4% de probabilidad). En Francia e Italia desaparece antes que en el resto de Europa, mientras surgen Chatelperroniense y Uluzziense, respectivamente.
- La desaparición de los neandertales tuvo lugar en diferentes momentos en diferentes regiones.
- El rango del Uluzziense quedó establecido en 46,45-39,2 ka cal BP. Al comparar este dato con los obtenidos para el Musteriense y el Chatelperroniense, se observa un solapamiento temporal entre los dos grupos humanos y las tres industrias de ca 5,4-2,6 ka (95,4% de probabilidad).
- Musteriense, Chatelperroniense y Uluzziense desaparecen al mismo tiempo.
Según Trinkaus (2011) los primeros restos de sapiens en Europa, muestran una sorprendente abundancia de rasgos neandertales y arcaicos perdidos o muy raros en los sapiens tempranos africanos. Estos rasgos no aparecen juntos, sino que se muestran o no en los ejemplares de una forma aleatoria, tal como cabría esperar en dos poblaciones morfológicamente distintas que se mezclan durante algunos milenios. Además, permanecen durante miles de años, hasta el gravetiense.
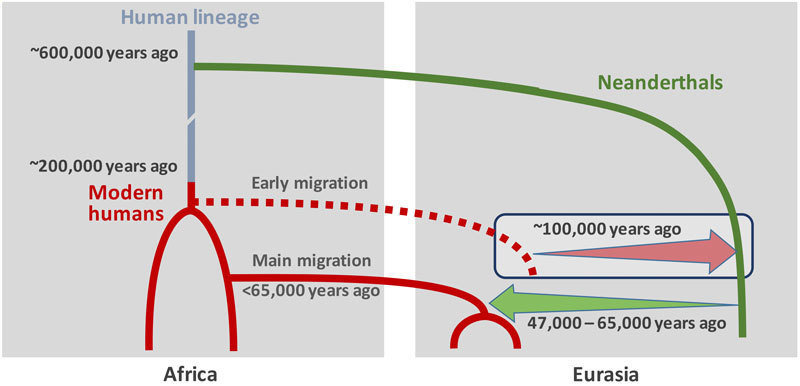
Según Jean-Jacques Hublin (2013; Hublin, 2014), se ha especulado con una llegada temprana del HAM a Europa conectada con los espectaculares cambios ambientales en MIS 3. El Protoauriñaciense se ha encontrado en la estratigrafía bajo las tefras Campanian Ignimbrite en Italia (Giaccio, 2005; Lowe et al 2012). Müller et al (2011), sostienen que la primera llegada del HAM pudo haber ocurrido bajo las condiciones suaves de GIS 14-13, momento que coincide con la aparición del Bohuniciense. Más tarde, el evento H 5 representó un importante deterioro climático hace ca. 48 ka y probablemente creó un vacío demográfico en grandes áreas que facilitaría la posterior expansión del HAM por los refugios del sur de Europa durante GIS 12, ca. 47 ka calibrado. El evento H 4 pudo haber retrasado la expansión del HAM solo en algunas áreas de Europa Occidental. La llegada del HAM a Europa antes de los albores del Auriñaciense (Willendorf II se ha datado en 43,5 ka; Philip R. Nigst et al, 2014) y un modelo escalonado para la colonización de algunas partes del continente tiene implicaciones importantes en cuanto a la evolución biológica y cultural en la zona y en las posibles interacciones con los neandertales. Explicaría no solo el Chatelperroniense, sino también los cambios culturales observados en el Musteriense de Tradición Achelense en el que aparecen pigmentos. La influencia habría llegado a los neandertales de forma directa por asentamientos a corta distancia y por difusión dentro de los grupos neandertales desde las zonas de contacto iniciales. La persistencia de rasgos arcaicos en los primeros HAM europeos se ha interpretado como señal de flujo de genes con la población neandertal (Zilhao, 2006); el análisis genético de Oase 1 (Qiaomei Fu et al, 2015) apoya esta teoría. Por otra parte, estos rasgos arcaicos se pueden observar en fósiles de HAM del Magreb y de otros lugares. Los europeos actuales muestran un bajo grado de introgresión neandertal, por lo que aquellas poblaciones tuvieron que ser sustituidas por otros grupos sin hibridación.
Se especula si el tiempo de contacto entre HAM y neandertales fue suficiente para permitir la transmisión de comportamientos culturales y simbólicos.
 |
| Escenarios de hibridación entre neandertales y sapiens. Ilan Gronau. |
- Los neandertales son una especie anterior a Homo sapiens y ancestral de ésta. Es una idea de principios del siglo XX a la que se ha adherido Wolpoff si bien considera que forman una única especie.
- La presencia de restos neandertales clásicos en época tardía es un argumento en contra. Podría aducirse que el salto evolutivo se hizo de forma local – en Próximo Oriente- mientras que en otras partes se mantenía la especie neandertal.
- Los neandertales son una subespecie del Homo sapiens, Homo sapiens neanderthalensis (B. G. Campbell, 1963) que, contribuyó con su acervo genético a la aparición del Homo sapiens sapiens (hipótesis de la evolución multirregional).
- Para Trinkaus, de acuerdo con las evidencias fósiles de Próximo Oriente, sería más que probable. Por otra parte, los rasgos en mosaico de los sapiens europeos hasta el gravetiense, implican hibridación. Considera además que los rasgos de conducta entre neandertales y humanos estaban lo bastante próximos entre sí como para permitir el intercambio genético.
- John Hawks (2012) proporciona una explicación adicional: el hecho de que los neandertales del musteriense ofrezcan unos rasgos clásicos mientras que en los últimos neandertales aparezcan rasgos en mosaico, podría deberse a la sustitución parcial de la población neandertal europea por otra población neandertal de origen asiático.
- Son dos especies diferentes con los neandertales formando una rama lateral que no guarda relaciones de ancestro con Homo sapiens. De hecho éste podría ser incluso más antiguo, ya que el antecesor común más reciente de neandertales y sapiens, según evidencias genéticas, vivió hace 550-690 ka (McDougall et al, 2005).
- Es la postura de Stringer y Gamble. También de Wynn, Overmann y Coolidge (2016). (Sustitución brusca de las poblaciones anteriores de neandertales por seres humanos de aspecto moderno. Hipótesis del reemplazo).
Pinchar aquí para repasar el concepto de especie.
- En el cromosoma X de los humanos actuales no-africanos, no se han detectado fragmentos arcaicos (por ejemplo, Skov et al, 2020). Tampoco se han detectado en el ADN mitocondrial. Esto sugiere que la descendencia de hembras neandertales con machos sapiens era inviable o estéril. Mason y Short (2011) propusieron que la introgresión neandertal pudo seguir la regla de Haldane: las interacciones genéticas interespecíficas producen esterilidad unisexual en la descendencia híbrida (Haldane, 1922). Esto se ha observado en insectos, aves y mamíferos: en algunas especies, los híbridos femeninos no se reproducen en absoluto o lo hacen con poca frecuencia y en otras el síndrome afecta a la descendencia masculina (Coyne, 1985; Haldane, 1922). Han sugerido que los únicos híbridos fértiles eran crías hembras con padres neanderthalensis y madres sapiens.
- La esterilidad en los mamíferos híbridos surge después de un tiempo de separación de las especies de al menos unos 2 Ma (Fitzpatrick, 2004;Wu, 1992). Por lo tanto, la hipótesis de Mason y Short requeriría una aparición de la esterilidad particularmente rápida, por lo que es poco probable. Sin embargo, pudo ser suficiente tiempo para unas barreras reproductivas parciales o incompletas, especialmente por mecanismos precigóticos, que requieren menos tiempo de desarrollo. Esto es consistente con una direccionalidad de la introgresión a través de algún desnivel de efecto.
- Para Armando Neves y Maurizio Serva (2012) la cantidad de ADN que compartimos con los neandertales es compatible con hibridaciones muy ocasionales (un par de individuos cada 77 generaciones). Estos raros cruzamientos son también compatibles con la ausencia de genes neandertales en el ADN-mt y en el Y-ADN. Sugirieron que H. neanderthalensis y Homo sapiens se aislaron reproductivamente a través de las diferencias culturales o sociales, y Schillachi y Froehlich (2001), que las dos especies podrían haber tenido problemas para reconocerse recíprocamente como compañeros adecuados. Currat y Excoffier (2011) sugieren una baja fertilidad de los híbridos; partiendo de la direccionalidad de la introgresión, consideran que la fertilidad híbrida puedo haber sido dependiente del contexto.
- La violencia, la xenofobia, el comportamiento de aprovisionamiento, las diferencias ontogenéticas, morfológicas y de comportamiento, que afectan al reconocimiento de parientes y para el apareamiento, el infanticidio y la selección sexual pueden haber sesgado la ocurrencia de entrecruzamiento o la supervivencia y la reproducción de los híbridos de una manera que podría explicar al menos parcialmente, el patrón de introgresión.
- Karenleigh A. Overmann y Frederick L. Coolidge (2013) examinaronn la hipótesis de un fuerte aislamiento reproductivo entre Homo neanderthalensis y el Homo sapiens, tomando como referencia la hibridación chimpancé-bonobo y la de los babuinos.
- En la hibridación de primates se ha observado aislamiento reproductivo precigótico, aislamiento reproductivo postcigótico con flujo de genes asimétrico y aptitud híbrida inferior (Curnoe et al, 2006; Aguiar et al, 2008; Stevison y Kohn, 2009), con el comportamiento (por ejemplo, filopatría; dispersión) y la estructura del grupo social influyendo en el flujo de genes y la estructura de la población (Tung et al, 2010). Si H. neanderthalensis y Homo sapiens se aislaron reproductivamente, pudo haber sido a través de comportamientos que impidieron la ocurrencia de apareamiento, por medio de una menor aptitud de los híbridos o por ambas razones.
- Pan troglodytes y P. paniscus son nuestros parientes más cercanos genéticamente (Green et al, 2010). La pequeña distancia genética entre ellos sugiere que podrían ser considerados como una sola especie, como se ha sugerido para H. sapiens y H. neanderthalensis. La similitud genética, sugiere que la hibridación habría sido probable para ambos pares de especies, dada la proximidad geográfica. De hecho, chimpancés y bonobos se cruzan en cautividad y producen híbridos aparentemente normales (Pan paniscus, P. t. schweinfurthii, P. t. troglodites , P. t. vellurosus y P. t. verus en todas las combinaciones; Curnoe y Thorne, 2003) posiblemente fértiles. Estos híbridos se caracterizan por ser intermedios en su morfología y comportamiento (Vervaecke , comunicación personal) y su vitalidad es normal.
- Los plazos y las razones de la especiación son paralelos entre los dos pares de especies: La separación de chimpancés y bonobos se produjo hace menos de 1 Ma, comparable al tiempo de separación entre neandertales y los primeros humanos modernos (Green et al, 2006, 2010; Hey, 2009). Ambos eventos de especiación pudieron haber ocurrido por aislamiento geográfico. Las diferencias morfológicas entre chimpancés y bonobos son sutiles; los bonobos, quizás más pedomórficos que los chimpancés. Los H. sapiens son igualmente pedomórficos en relación con H. neanderthalensis. Las distinciones de comportamiento entre los chimpancés y los bonobos incluyen la cooperación, los patrones de dispersión, la dominación, las estructuras sociales, el aprovisionamiento, la territorialidad, la tolerancia y el apareamiento (Hare et al, 2007; Parish y de Waal, 2006; Prüfer et al, 2012). H. neanderthalensis y Homo sapiens también pueden haber diferido en comportamientos tales como el aprovisionamiento, la dieta, la producción de herramientas, el arte y la creatividad, el lenguaje, la espacialidad y el tamaño del grupo (Kuhn y Stiner, 2006; Lalueza-Fox et al, 2011; Richards et al, 2009; Spikins et al, 2010; Wynn y Coolidge, 2012, Bruner, 2004, 2010; Bruner et al, 2003; Gunz et al, 201; Lieberman et al, 2002; Pearce et al, 2013; Paixão-Cortés et al, 2013; Traynor et al, 2015).
- El aislamiento reproductivo entre neandertales y HAM pudo mostrar las siguientes dimensiones:
- La violencia intergrupal y la xenofobia Neandertal podrían haber reducido la probabilidad de cruzamiento.
- Las diferencias en el comportamiento de aprovisionamiento podrían haber aumentado la probabilidad de cruce entre machos neanderthalensis y sapiens hembras.
- Las diferencias morfológicas entre las dos especies humanas observadas en los fósiles pudieron haber influido en el reconocimiento de pareja mediante factores de desarrollo ontogenético y selección sexual. Estas diferencias pueden haber proporcionado ventajas a los varones neanderthalensis.
- Los neandertales muestran un desarrollo dental más temprano por lo que alcanzaban el reconocimiento de pareja y la maduración a una menor edad.
- El HAM era pedomórfico en relación con el neandertal, que se caracteriza por una mayor masa muscular (Ben- Itzhak et al, 2005; Gunz et al, 2010; Lieberman et al, 2007; Smith et al, 2010). Estas diferencias fisiológicas podrían haber influido especialmente en la elección femenina. Las hembras neandertales podrían haber juzgado a los varones HAM como una pareja inadecuada. Por el contrario, para las hembras sapiens los varones neandertales, debido a su mayor robustez, pudieron haber sido atractivos. Este patrón de preferencia de machos más robustos de otra especie se observa en varias hibridaciones de primates.
- Posiblemente los machos híbridos tuviesen ventaja entre los HAM, pero desventaja entre los neandertales.
- La paternidad incierta, el comportamiento intermedio y el reconocimiento de parentesco debilitado pudieron haber reducido la supervivencia de los híbridos.
- En definitiva, el patrón de introgresión detectado no pudo ser producido por la esterilidad o la inviabilidad de los híbridos debido a la cantidad insuficiente de tiempo entre la separación y el reencuentro de las dos especies. Por el contrario, la direccionalidad de la introgresión pudo haber sido influenciada por formas de aislamiento reproductivo típicas en los primates, como el aislamiento precigótico y la supervivencia y reproducción postcigótica diferenciales.
 |
Comparación de la cronoestratigrafía de Willendorf II (AH 3) y las dataciones de los últimos neandertales datados directamente. Philip R. Nigst et al (2014).
|
Enlaces:
The Neandertal genome paper: Green, R.E., et al (2010). A Draft Sequence of the Neandertal Genome. Science 328:710.
Ofer Bar-Yosef. 2013. Neanderthals and Modern Humans Across Eurasia.
Genéticamente no hay rastro de descendencia por via paterna de los neandertal. Adn mitocondrial cromosoma y perdido. Por ello es mas lógico pensar que las crias de homo sapiens macho y neandertal hembra fueron los únicos supervivientes. Tal vez porque la hembra neandertal es mas robusta.
ResponderEliminarEn el cromosoma X de los humanos actuales no-africanos, no se han detectado fragmentos arcaicos (por ejemplo, Skov et al, 2020). Tampoco se han detectado en el ADN mitocondrial. Esto sugiere que la descendia de hembras neandertales con machos sapiens era inviable o estéril.
Eliminar