 |
| Posición filogenética de LD 350-1. Emily M. Eng, NG Staff. |
Colaboradores de Kimbel descubrieron en 1994 un
maxilar superior casi completo en Hadar, de antigüedad superior a 2,3 Ma, que
confirmaba la separación de linajes en aquella fecha. Los estudios
paleoambientales indican que la región de Hadar albergó extensiones abiertas de
sabana en el Plioceno tardío. Ambos datos concuerdan con el modelo del género Homo como resultado de la adaptación a
los espacios abiertos a través del uso de herramientas.
Al igual que los Australopithecus y Paranthropus
los Homo rudolfensis y habilis se desarrollan
en el Este y Sur de África y allí permanecen hasta su extinción, sustitución o dilución
genética en sus descendientes: el grado erectus (Homo erectus/ergaster).
Se han propuesto como primeros ejemplares correspondientes
al género Homo algunos fósiles
datados entre 2,8-2,3 Ma.
 |
| LD 350-1 |
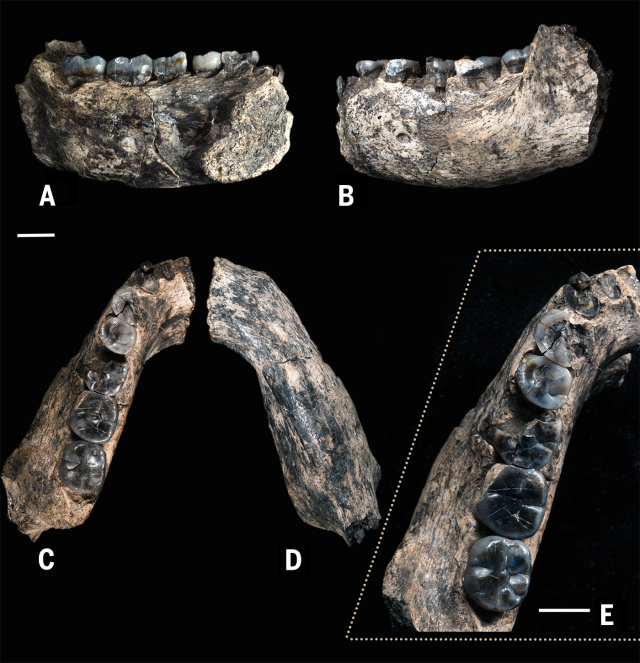
LD 350-1
Hallado en 2013 en Ledi-Geraru, Afar, Etiopía, una mandíbula izquierda parcial con dientes datada en 2,80-2,75 Ma. Este espécimen combina rasgos primitivos presentes en Australopithecus con morfología derivada observada los Homo posteriores (Brian Villmoare et al, 2015).- LD 350-1 está compuesta por dos piezas separadas por una fractura vertical al nivel de M1. Están asociadas algunas coronas dentales y fragmentos de raíz que se hallaron sueltos. Los molares son estrechos y los premolares simétricos como en los Homo posteriores.
- Se diferencia de Australopithecus garhi en el inferior tamaño de los dientes. El área del M2 inferior de LD 350-1 es un 64,7% de la de BOU-VP-12/130.
- La morfología dental y mandibular es más derivada que la de Australopithecus sediba.
- P4 y M3 son más pequeños que los de la mandíbula KNM-WT 8556, de asignación incierta y datada en ca 3,3 Ma.
- LD 350-1 tiene una morfología más primitiva que habilis o rudolfensis en la porción anterior del corpus mandibular si bien es similar a habilis en otras características como la curvatura de la fila de dentición anterior.
LD 350-1 podría ser un miembro final de Australopithecus afarensis, dado un número de similitudes morfológicas, como las dimensiones generales de la dentición y corpus mandibular y la orientación de la sínfisis mandibular, pero otras características, como la ausencia de patrones distintivos de desgaste entre el canino superior y el P3 mandibular y la profundidad uniforme del corpus mandibular entre el P3 y el M2, no son compatibles con afarensis.
 |
| Reconstrucción especular de LD 350-1. Barra = 1 cm. |
Según Erin N. DiMaggio et al (2015), la fauna asociada sugiere un hábitat abierto o mezcla de praderas mixtas y matorrales con bosques de galería bordeando ríos o humedales. El paisaje era probablemente similar al actual Serengeti o Kalahari.
Este hábitat es consistente con la hipótesis de que los cambios en el medio forzaron la evolución humana.
El fósil demuestra que algunos de los cambios dentales y mandibulares entre Homo y Australopithecus ocurrieron temprano y puede indicar una divergencia importante en adaptaciones dietéticas..
La datación de LD 350-1 y la fauna asociada se ha realizado triangulando varios métodos, la datación de capas de cenizas volcánicas mediante argón 40 / argón 39, geoquímica, paleomagnetismo y fauna. La datación ofrece ciertas dudas debido a que los fósiles se hallaron fuera de contexto.
 |
| Posición cronológica de LD 350-1. Nature. |
Sts 19
Sterkfontein, miembro 4, Sudáfrica. 2.7-2.5 Ma?. Ha sido
imposible obtener una datación de total certeza.
Base del cráneo bien conservada hallada en los escombros de
una explotación minera. Broom et al (1950) quedaron impresionados por su
morfología parecida a la humana, y, más tarde, Clarke (1977) consideró que
debía atribuirse a Homo sp. en lugar
de A. africanus. Esta posibilidad fue
rechazada por Dean y Wood (1982) a favor de una amplia gama de variación de A. africanus, pero Kimbel y Rak (1993)
propusieron su asignación a Homo.
El hueso temporal de Sts 19, muestra una posición semejante a la humana. La forma del tímpano y los elementos petrosos y sus relaciones con la anatomía craneoespinal adyacente, no se observan en ningún otro cráneo de Sterkfontein, pero son cualitativamente similares a los de algunos australopitecinos robustos. La anatomía de la base del cráneo es menos derivada que la observada en los fósiles de Homos de más de 2 Ma (por ejemplo, OH 13, OH 16, OH 24, KNM-ER 3891,Omo L. 894-1, STW 53), en los que la fosa mandibular se comprime anteroposteriormente, y el plano preglenoideo es muy empinado.
KNM-BC 1
Formación Chemeron, Kenia, ca. 2,4 Ma. 1966.Hill et all 1992; Sherwood et al, 2002 consideraron este hueso tempral fragmentario como el ejemplo más antiguo del linaje Homo basándose principalmente en la posición medial de la fosa de la mandíbula en relación con la pared lateral de la caja craneana, que pensaban que reflejaba la expansión del cerebro, así como en varias características cualitativas de la región glenoidea y el elemento petroso.
Se ha cuestionado la importancia filogenética de estas características ya que su presencia y expresión varían ampliamente entre los taxones de homínidos fósiles y/o pueden ser interpretadas como simplesiomorfias.
Las morfometrías exhibidas muestran afinidades tanto con Homo como con Australopithecus garhi (Asfaw et al 1999).
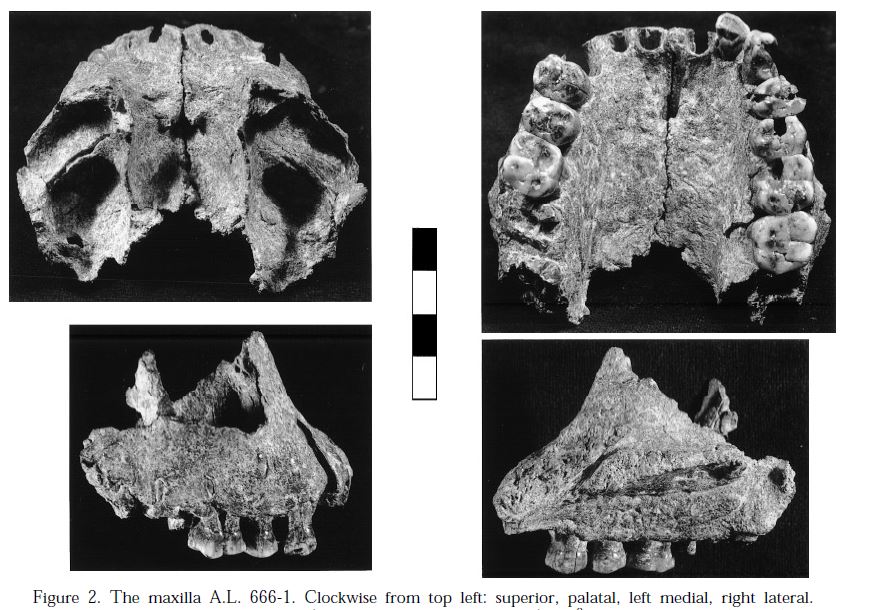
AL 666-1
Formación Hadar, Etiopía, ca. 2,3 Ma. Kimbel et al (1996).Maxilar fragmentado en 35 piezas, muy probablemente de un individuo de sexo masculino, con la mayor parte de la dentición.
Similar a los fósiles de Homo habilis KNM-ER 1813, L 894-1, y a los hallazgos de Bed 1 y Bed 2 inferior de la Garganta de Olduvai en su prognatismo subnasal bajo, procesos frontales evertidos, paladar relativamente amplio, coronas simétricas de los caninos superiores, alto ratio del área de la corona de P3/P4 superiores, M1 superior mesiodistalmente alargado y contorno oclusal romboidal del M2 superior
Carece de la morfología derivada cigomáticoalveolar, infraorbitaria y subnasal de KNM-ER 1470 (Homo rudolfensis, Kimbel et al, 1997).
Sin embargo, el P4 superior de AL 666-1 es vestibulolingualmente amplio en comparación con ejemplares más recientes, similar a los premolares de homínidos no robustos de los niveles anteriores a 2.0 Ma en la Formación Shungura, Etiopía (Suwa, 1990).
 |
| AL 666-1 |
Huesos del pie de Omo (Formación Shungura)
Astrágalo (Omo 323-76-989; Miembro G; Localidad 323) y calcáneo (Omo 33-74-896; Miembro E; Localidad 33) datados respectivamente en 2,2 y 2,36 Ma (Gebo y Schwartz, 2006). El astrágalo es similar en longitud al de los humanos modernos y a KNM-ER 813, con una tróclea estrecha, un cuello corto y una cabeza estrecha tanto en términos absolutos como relativos. El calcáneo posee un gran y robusto tubérculo peroneo; es ligeramente más corto que el de los humanos modernos; estrecho, con un sustentáculo del astrágalo estrecho en comparación con la población de la Sima de los Huesos y los neandertales.
Podrían pertenecer a Homo o a Paranthropus (Adrián Pablos, 2015).
Dientes
Dientes
- Miembros E-G, Formación Shungura, Etiopía, ca. 2.4-2.0 Ma. Suwa, White y Howell, 1996. Estos dientes muestran características derivadas observadas solo en restos de más de 2,0 Ma de H. habilis y rudolfensis (Suwa et al, 1996). M1 inferiores son bucolingualmente estrechos, mientras que las coronas de los P3 inferiores son igualmente estrechas y muestran una menor asimetría de la línea oclusal y un borde marginal mesial más desarrollado que en la mayoría de los homólogos de los primeros australopitecinos (A. afarensis y A. africanus). Suwa et al. (1996) señalaron que las coronas de los P3-M1 inferiores son relativamente grandes, con talónidos premolares expandidos, semejantes a los de KNM-ER 1802 (Wood, 1991) y UR 501 (Bromage et al 1995) (Homo rudolfensis). Se ha sugerido que este patrón dental puede haber sido primitivo para el linaje Homo.
- KNM-WT 42718 (Formación Nachukui, West Turkana, 2,3 Ma; Prat et al, 2005). M1 con una corona más pequeña, similar, sobre todo en su tamaño y su alto grado de compresión bucolingual, a los dientes de la Garganta de Olduvai y Koobi Fora atribuidos a Homo habilis (Prat et al., 2005). Combinado con la morfología de AL 666-1, esta evidencia apunta a una divergencia de los morfotipos de los Homo anteriores a 2,3 Ma.
- MLP-1549. Mille-Logya, Afar, Etiopia. Corona de un segundo molar superior. 2,5-2,4 Ma. Zeresenay Alemseged et al, 2020.
Te puede interesar:
|
No hay comentarios:
Publicar un comentario