|
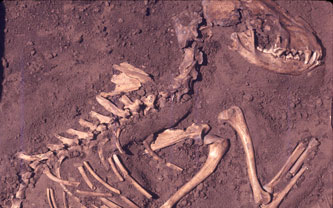
| Primeros Hominini. Haile-Selassie, Melillo y Su, 2016. |
La especie más conocida es Australopithecus afarensis (3.7 a 2.9 Ma) recuperado en gran medida en Tanzania (Laetoli) y Etiopía (Hadar, Dikika, Woranso-Mille), con un gran número de fósiles que incluyen esqueletos parciales y cráneos relativamente completos. La evidencia fósil disponible sugiere que Au. afarensis es un descendiente directo de Au. Anamensis, el mejor ejemplo de gradualismo filogenético dentro de los Hominini tempranos.
- La diferencia entre anamensis y afarensis parece una cuestión de grado de consecución de las mismas adaptaciones. Este hecho, junto con la similitud de contextos ecológicos de los yacimientos y la falta de solapamiento en las dataciones, nos hace pensar que se trata de cronoespecies o paleoespecies dentro de un proceso de anagénesis, descartando una cladogénesis.
En este caso, Australopithecus africanus sería el taxón hermano de estas especies.· Si ordenamos los mejores ejemplos de ambas especies en una secuencia Kanapoi→Allia Bay→Woranso Mille→Laetoli→Hadar, surge de inmediato la idea de una anagénesis.· Los fósiles de Woranso Mille muestran un mosaico de rasgos como-anamensis y como-afarensis (Haile-Selassie, 2010).Queda descartada por tanto la hipótesis de M. G. Leakey de que Ardipithecus. ramidus y Australopithecus afarensis forman ramas laterales en la evolución humana, mientras que Australopithecus anamensis es un buen candidato para formar parte de nuestros ancestros. O afarensis y anamensis forman parte de nuestros ancestros o ninguno de ellos.
El descubrimiento de un fragmento de mandíbula en África Central (Australopithecus bahrelghazali), a unos 2.500 km de los yacimientos de afarensis, fue el primer indicio de la presencia de más de una especie de Hominini y de una amplia distribución geográfica. La cuestión se reavivó con el descubrimiento en Kenia de fósiles asignados a Kenyanthropus platyops y del ejemplar BRT-VP-2/73, un pie parcial (el Pie de Burtele) datado en 3,4-3,3 Ma, hallado en Woranso-Mille, Afar, Etiopía, con un hallux oponible y una adaptación locomotora que lo diferencia de Au. afarensis y lo acerca a Ar. ramidus. Recientemente, el registro arqueológico se enriqueció con el hallazgo de Au. deyiremeda en Woranso-Mille, diferente de afarensis en la morfología dental y mandibular y de platyops en el maxilar. Los sedimentos de origen son contemporáneos de los de Maka y Dikika donde aparecieron restos de afarensis. No se puede descartar que BRT-VP-2/73 pertenezca a deyiremeda.
Salvo en el caso de afarensis, la validez de estos taxones ha sido cuestionada, por el pequeño tamaño de la muestra y la falta de evidencia de diferencias de nicho ecológico, si bien la composición isotópica y el espesor del esmalte dental sugieren diferencias dietéticas entre Au. bahrelghazali y Au. afarensis. BRT-VP-2/73 demuestra claramente la existencia de un nicho más arbóreo, frente al nicho más terrestre de Au. afarensis.
- En Woranso-Mille, afarensis y deyiremeda parecen haber estado viviendo en simpatría directa. Ambas especies podrían haber sido generalistas con una de ellas abarcando un nicho más amplio o con diferentes estrategias en alimentos de reserva, tal y como hoy sucede con los chimpancés y gorilas que comparten áreas en África Ecuatorial. Australopithecus afarensis, estaba más adaptado a ecosistemas relativamente abiertos mientras que Australopithecus deyiremeda conserva adaptaciones a la vida arbórea.
Las relaciones filogenéticas entre estas especies son difíciles de determinar:
- En Au. bahrelghazali, K. platyops, y Au. deyiremeda son evidentes las características dentales derivadas, pero ninguna de ellas presenta el conjunto completo de sinapomorfías que caracterizan Paranthropus u Homo. Con respecto a afarensis son más derivados en algunas características y mas primitivos en otras.
- La morfología dental de Au. afarensis es intermedia entre las de Ardipithecus y Au. Anamensis por un lado y la de Paranthropus por otro. Podría representar la condición plesiomórfica con relación a todos los Hominini posteriores a 3 Ma, pero esto desafía el punto de vista tradicional de que afarensis es el antepasado de todos ellos. Para Richard Leakey, afarensis no es un ascendiente de Homo. Por otra parte el espesor del esmalte y tamaño de sus dientes (grandes) son completamente diferentes a los que aparecen en humanos y chimpancés (pequeños).
- Algunos autores se han fijado en las características robustas de AL 442-2 para presentar a Australopithecus afarensis como un antecesor de los parántropos. Si esto es así, la separación de los linajes grácil y robusto estaría ya en curso en estos primeros individuos.
- Es controvertida la vinculación de la morfología craneodental de K. platyops con la de Homo rudolfensis.
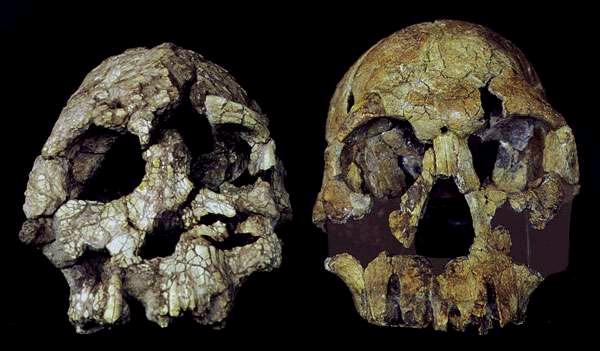 |
| Kenyanthropus platyops (izquierda) y Homo rudolfensis (derecha) |
Te puede interesar:
|