(A) Paleolítico Superior Inicial (IUP), Paleolítico Medio (MP), y yacimientos arqueológicos de Modo 1 (core and flakes) y con restos humanos en el centro y norte de Asia.
(B) Ruta sugerida por Goebel (2015) y yacimientos del IUP.
1. Anghilak cave, 2. Teshik Tash, 3. Khudji, 4. Obi-Rakhmat, 5. Shugnou, 6. Chagyrskaya, 7. Okladnikov, 8. Denisova, 9. Ust-Karakol, 10. Kara-Tenesh, 11. Kara-Bom, 12. Luotuoshi, 13. Tongtian Cave, 14. Gouxi, 15. Lenghu, 16. Heimahe 1, 17. Chikhen Agui, 18. Tsagaan Agui, 19. Tolbor 4, 20. Kharganyn Gol 5, 21. Orkhon 1 & 7, 22. Makarovo 4, 23. Kandabaevo, 24. Varvarina Gora, 25. Tolbaga, 26. Temple Canyon 1, 27. Shuidonggou 1, 27. Shuidonggou 2, 27. Shuidonggou 7, 28. Shuidonggou 9, 29. Fanjiagouwan (Salawusu), 29. Yangshugouwan (Salawusu), 30. TX08, 31. TX03, 32. Liujiacha, 33. GY03, 34. ZS08, 35. ZL05, 36. Shuangbuzi, 37. Shixiakou 2, 38. Xujiacheng, 39. Changweigou, 40. Gutougou, 41. Wulanmulun, 42. Yushuwan, 43. Shiyu, 44. Dingcun (7701), 45. Licunxigou, 46. Fuyihe (Xiachuan), 47. Beiyao, 48. Zhiji, 49. Tashuihe, 50. Laonainaimiao, 51. Huangdikou, 52. Longquandong, 53. Xiaonanhai, 54. Dangcheng (Shidie), 55. Xibamaying, 56. Jinsitai, 57. Tianyuandong, 58. Upper cave, 59. Wangfujing, 60. Zhuacun, 61. Huangniliang, 62. Dazhushan, 63. Xiaogushan, 64. Miaohoushan, 65. Xianrendong, 66. Shimenshan, 67. Zhoujiayoufang, 68. Yanjiagang, 69. Guxiangtun, 70. Shibazhan (75075), 71. Salkhit, 72. Ust’-Ishim.
|
 |
| Dientes de Daoxian. (María Martinón-Torres et al, 2017). |
La historia demográfica de Asia oriental no parece lineal y sugiere profundas subdivisiones geográficas y subestructuras de la población, tal vez similar a lo que se identifica en África. Las investigaciones genéticas han revelado episodios de hibridación entre HAM y los anteriores pobladores de Asia. Este intercambio genético agrega un importante factor que contribuye a la variabilidad morfológica de los grupos humanos. (María Martinón-Torres et al, 2017).
Durante muchas décadas, erectus se ha utilizado como un término general para referirse a casi cualquier Homo encontrado en Asia durante el Pleistoceno hasta la aparición del Homo sapiens (Xing, Martinón Torres y Bermúdez de Castro, 2018), pero estudios recientes sugieren que se ha simplificado en exceso y que pueden pertenecer a varios linajes diferentes.
- Algunos fósiles presentan algunas características primitivas en relación a erectus: el endocast de Maba es estrecho en los lóbulos frontales y corto y aplanado en las áreas parietales; el neurocraneo de Xuchang es bajo e inferiormente amplio; la muestra de Xujiayao comprende un cráneo grueso y fuerte y molares grandes y complejos; el incisivo central superior de Panxian Dadong muestra proyecciones conspicuas en su superficie lingual.
- Por otra parte, muestran también características derivadas en la línea del sapiens: el agrandamiento y la dilatación neurocraneal en el espécimen de Xuchang; una escama temporal alta y redondeada, superficies bucales lisas y oclusales simplificadas de los premolares superiores, y un contorno simétrico de la corona con una cúspide lingual pronunciadamente reducida de P3 en la muestra de Xujiayao; posiciones derivadas de los lóbulos frontales en relación con las órbitas y morfologías del seno frontal y la escama frontal en Maba; y la forma moderna del contorno de la corona similar a la humana de los premolares superiores e inferiores y del canino inferior similar a un incisivo en Panxian Dadong.
- Además, Xuchang y Xujiayao tienen algunas características que se encontraron de forma clásica en el linaje neandertal. Xuchang muestra una fóvea suprainácea y su toro nucal y el laberinto temporal han sido descritos como de tipo neandertal. De manera similar, las morfologías del laberinto temporal y mandíbula de Xujiayao también se parecen a las de los neandertales.
- Fósiles como Dali y Yunxian han sido clasificados como "arcaico" o "post-erectus" y posibles representantes del taxón H. heidelbergensis. Estos, junto con el hipodoma Xujiayao o Xuchang, podrían ser candidatos potenciales para representar a los Denisovanos.
Dos teorías se disputan la interpretación de los restos arqueológicos:
- Ocupación temprana: Interpreta como restos de HAM algunos hallazgos en China datados en 100 ka o incluso antes.
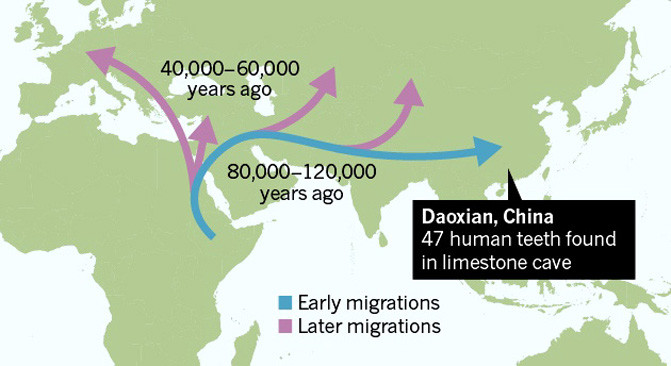
- Los dientes de la Cueva Fuyan (Daoxian), datados entre hace 120-80 ka (Wu Liu et al, 2015). 47 dientes humanos que pueden atribuirse inequívocamente a H. sapiens. Son más derivados que los de otras poblaciones contemporáneas de África y Europa, incluyendo Qafzeh y más cercanos en morfología a los humanos modernos contemporáneos y a muestras del Pleistoceno Tardío Tardío. Estas diferencias apuntan a la posibilidad de que el sapiens del Levante Mediterráneo y el de China, representen dos demes diferentes (María Martinón-Torres et al, 2017).
- Aunque corresponden a un período en el son infrecuentes, los dientes de Daoxian presentan caries, y eso podría arrojar dudas sobre su antigüedad. Podría significar que los primeros HAM que llegaron a Asia eran más propensos a desarrollar caries o podría reflejar un cambio en la dieta. Esto es particularmente importante a la luz de los estudios que sugieren que los HAM pudieron haber explotado los recursos de la selva tropical en el sur y sureste de Asia antes de lo que generalmente se supone (por ejemplo, Demeter et al, 2012; Roberts et al, 2015 , 2016; Westaway et al, 2007).
 |
| Dientes de Panxian Dadong. María Martinón-Torres et al, 2016. |
- Wu Liu el al (2013), han estudiado cuatro dientes de homínidos hallados en Panxian Dadong (Sur de China), datados en 300-130 ka. Los dientes muestran un mosaico de rasgos primitivos y derivados que los alinean con los del Pleistoceno Medio y Superior de Asia y Europa. Están ausentes los rasgos típicos de los neandertales. Sin embargo, los rasgos derivados no constituyen un diagnóstico suficiente para relacionarlos con Homo sapiens. El estatus derivado con respecto a otras muestras (Hexian, Chaoxian, Zhoukoudian y Sangiran) clasificadas como erectus (pero no con respecto a las de Kabuh Formation analizadas por Zanolli, 2015) sugieren que Panxian Dadong podría representar un demo diferente.
- Dos dientes permanentes de homínido recuperados in-situ en Lunadong, Guangxi Zhuang, China se han asignado a HAM (Christopher J. Bae et al 2014). Según múltiples fechas de series de uranio, se pueden fechar con seguridad entre 126,9 ± 1,5 ka y 70,2 ± 1,4 ka. Presentan características típicas de los HAM, pero las raíces son robustas, lo que no permite una asignación rotunda (María Martinón-Torres et al, 2017).
- Ganqian 220-94, Shen et al, 2002.
 |
| Dientes de Xujiayao. María Martinón Torres et al, 2016. |
- Xujiayao-Houjiayao (XH), en el noroeste la cuenca del Nihewan, norte de China, excavado en 1976, 1977 y 1979. Dientes y fragmentos óseos, con características típicas de una forma no neandertal temprana de humanos arcaicos finales.
- Varias bóvedas craneanas (15 fragmentos), dos mandíbulas y tres dientes aislados de un mínimo de diez individuos Homo arcaicos, además de 30.000 artefactos líticos y restos de fauna asociada. En 2007 y 2008, se excavó sistemáticamente la esquina noroeste del sitio.
- Un fragmento de maxilar izquierdo infantil consiste principalmente en la porción alveolar y tres dientes permanentes recientemente erupcionados: I1, C1 y M1, además de la raíz de un segundo molar temporal. I1 y C1 se distinguen por su forma de pala. Las dimensiones de dientes y paladar sugieren que perteneció a un varón. Los perikymata son más similares a los del HAM que a los de los neandertales (Xing et al, 2015). Presenta un defecto en el esmalte evidente en la superficie vestibular de los premolares y un surco profundo que rodea la corona, particularmente pronunciado en las partes lingual y distal del diente.
- Xujiayao 15 es un hueso temporal cuyo laberinto está dentro de la variación neandertal. No presenta ninguna otra morfología neandertal.
- El estudio de la muestra dental (Song Xing et al 2014), revela un mosaico de rasgos primitivos y derivados que se pueden resumir de la siguiente manera:
- Son diferentes a los de los humanos modernos y arcaicos recientes.
- Presentan algunas características que son comunes, pero no son exclusivas, en los neandertales.
- Conservan algunas conformaciones primitivas que aparecen en los homínidos del Pleistoceno Inferior y Medio de Asia Oriental.
- Por tanto, el estatus taxonómico de la población es incierto. Los resultados, sugieren la posibilidad de que un linaje homínido primitivo pudo haber sobrevivido hasta el Pleistoceno Superior en China.
- Ele hemimaxilar izquierdo de un juvenil Xujiallao 1 ha revelado una ontogenia dental equivalente a la de un humano monderno (Son Xing et al, 2019)
- Hua Tu et al (2015) han datado, por 26Al/10Se, dos muestras de cuarzo por debajo de la capa cultural más antigua, que han proporcionado una edad media de 240 ± 50 ka. Esta datación es consistente con otra reciente por post-IR IRSL (postinfrarrojos-luminiscencia estimulada por infrarrojos) que arrojó un rango de hace 224 ± 22 a 161 ± 13 ka para las capas con restos culturales. De acuerdo con la evidencia de fauna y palinológica que es indicativa de un período de clima frío, estos resultados apoyan la proposición de que el sitio XH debe corresponder a MIS 6 (ca 190-130 ka).
- Hong Ao el al (2017) han aplicado el método del ESR a granos de cuarzo del nivel donde aparecieron los restos fósiles, estableciendo una horquilla temporal entre 370-260 ka.
- María Martinón-Torres et al (2017), han sugerido que los fósiles de Xujiayao podrían ser buenos candidatos para representar a los denisovanos.
- Bailiandong (Guangxi, China). >160 ka , Guangxi. Shen et al, 2001
- Liujiang (Guangxi, China). 153-68 ka, muy probablemente entre 139-111 ka, Brown, 1992; Shen et al, 2002. Cartmill y Smith, 2009).
- Maba 1, Guangdong. Cráneo parcial, Hallado en 1958, con una datación mínima de ca 0,13 Ma por U-series y máxima de 0,3 Ma, con un boquete de 14 cm producto de un fuerte golpe con un objeto pesado romo. El herido de Maba tuvo que necesitar ayuda de su grupo social. Asociado a restos de mamíferos.
- Rasgos primitivos en general, pero también están presentes rasgos derivados y nuevos asociados con las relaciones espaciales de la cara. Se ha descrito una mezcla similar para la población de la Sima de los Huesos. La capacidad craneal de ca 1.300 cc, y las proporciones endocraneales están dentro de los rangos de los humanos modernos, los neandertales y heidelbergensis. Los lóbulos frontales son estrechos y las zonas parietales cortas y aplanadas, como en heidelbergensis y erectus. La posición de los lóbulos frontales en relación con las órbitas, la morfología del seno frontal y la curva de la escama frontal se deriva más, siendo similar a neandertales y HAM. Podría incluirse en el linaje neandertal (Pope, 1992) o representar un caso de convergencia (Wu y Bruner, 2016). Bermúdez de Castro y Martinón defienden un origen común de las poblaciones asiáticas y europeas que explicaría estas similitudes.
- Se le asocia con el cráneo de Hathnora, Narbada (India), datado hace más de 0,15 Ma.
- Recientemente se han descrito nuevos hallazgos y se ha efectuado una datación del travertino, pero el contexto no está claro y tampoco la relación de los fósiles con el travertino y entre sí (Dongfang Xiao et al, 2014).
- Zhirendong, Guangxi. Dos molares (probablemente M2 y M3) y un gran fragmento de mandíbula. 110-106 ka, Liu et al., 2010. Cai et al, 2016.
- La mandíbula presenta todos los elementos sinfísicos anteriores primarios de un mentón humano moderno, como un tubérculo proyectado sinfiseo, fosas mentales distintas, tubérculos laterales modestos y una sínfisis vertical (Liu et al, 2010), combinación que se considera típica y posiblemente exclusiva de H. sapiens. Sin embargo, la robustez del corpus ubica al fósil cerca de los humanos arcaicos del Pleistoceno Tardío. Este patrón en mosaico podría sugerir un flujo de genes entre H. sapiens y una población más arcaica (María Martinón-Torres et al, 2017). Otros investigadores han señalado la posibilidad de que la mandíbula Zhirendong represente un erectus grácil (Dennell 2010 , 2014), el surgimiento de una población con rasgos sapiens, de un modo similar a como debió de suceder en África (teoría multirregional).
- A pesar de su desgaste, en los molares es posible identificar características dentales claras correspondientes a H. sapiens y fuera de la variación de erectus (María Martinón-Torres et al, 2017).
- Cueva de Tontianyan, Liujiang, Guangxi-Zhung. Esqueleto incompleto de un joven adulto de 1,57 m, craneo completo con rasgos craneales muy modernos, fémures parecidos a los de neandertal o primitivos por lo grueso de la pared ósea y sin industria asociada. 139-111 ka.
- Xuchang 1 (Li et al, 2017), Hallado en 2007 cerca de Xuchangm Hunan por el equipo de Li Zhanyang. Fragmentado en 16 partes con membranas nerviosas fosilizadas y datado por termoluminiscencia entre 100-80 ka. 80.000 y 100.000 años. Asociados, se recuperaron algunos de huesos de animales modificados probablemente por herramientas. El análisis de las huellas de uso reveló que se trataba de instrumentos de hueso utilizados para la perforación y raspado de sustancias animales y que algunos pueden haber tenido mango durante el uso. En 2014 fue hallado otro cráneo, datado entre 125-100 ka. Los dos cráneos muestran características de erectus, como una bóveda neurocraneal baja, neurocráneo liso y mastoides cortos con inclinación hacia dentro, pero también de Homo sapiens. Además, presentan un hueso occipital y una estructura de laberinto de oído interno similar a neanderthalensis.
- Xiu-Jie Wu et al (2012) analizaron los fósiles Sangiran 4, Chaoxian 1, Xujiayao 1, y Changyang 1, encontrando un suelo nasal en dos niveles similar al que presentan los neandertales, mientras que los humanos modernos mostramos un solo nivel.
- Guanjun Shen et al (2013) han datado muestras de la cueva de Huanglong, norte de Hubei, China central, mediante espectrometría de masas de la serie de U, de alta precisión. En el yacimiento, se descubrieron siete dientes de Homo y docenas de herramientas de piedra y artefactos de hueso, junto con posibles sedimentos quemados. La morofología corresponde con la del HAM, si bien las raíces son robustas. En particular, Huanglong I presenta una forma de doble pala, una característica que es probablemente autapomórfica de H. sapiens (Liu et al, 2010; María Martinón-Torres et al, 2017).
 |
| Modelo de Continuidad con Hibridación. Características morfológicas que sugieren una aportación genética neandertal. |
- Existen diversas interpretaciones para estos fósiles de finales del Pleistoceno Medio:
- Son el resultado evolutivo del erectus antes de su extinción.
- Constituyen una posición intermedia entre erectus y sapiens, de acuerdo con el modelo Multirregional.
- Wu Xinzhi (1998) propuso el modelo de continuidad con hibridación: mosaico erectus y sapiens con flujo genético de neandertales. Las características morfológicas que denotan este flujo genético serían (ver figura más arriba):
- A. Órbita circular y margen orbital inferolateral agudo (Maba).
- B. Prognatismo (Najing 1).
- C. Moño occipital (Liujiang).
- D. Protuberancia de la superficie entre la apertura piriforme y la órbita (Dali).
- E. Orientación más lateral de la superficie anterolateral del proceso frontoesfenoidal del hueso cigomático (UC102).
- Para Liu et al (2003) y Zhang et al (2004), estas características no son evidencias suficientes para deducir una aportación genética neandertal.
- Pertenecen a poblaciones procedentes de África, relacionadas con Broken Hill, Bodo o Ndutu, que reemplazaron a erectus, tal vez con cruzamientos.
 |
| Olas de poblamiento Out of Africa del HAM. Los dientes de Lunadong han sido datados con seguridad entre 127-70 ka. |
 |
| Interpretación de los hallazgos de Daoxian. Nature. |
- Ocupación tardía. Aparición del HAM en África hace 190-160 ka y Out of Africa hace 50 ka.
- Un fémur incompleto hallado en 2008 por Nikolay Peristov en Ust-Ishim (Omsk) ha sido datado por radiocarbono en 45 ka. Se trata del resto de sapiens más antiguo datado directamente.
- Con menos de 60.000 años se han encontrado fósiles en Longtanshan, Salamushu, Shiyu, Zhiyang, Wushang, Shuicheng, Tianyuan Cave.
- En la Cueva Superior de Zhoukoudian (Upper Cave) se hallaron en 1933-1934 partes de tres esqueletos datados en 35,1-33,5 ka (Li et al, 2018).
- Con 30.000 años se han encontrado restos en Quiziang (o Yiyuan).
- Ziyang, 40-35 ka
- Tianyuan (Fangshan, norte de China), 42-39 ka. Shang et al, 2007; Cartmill y Smith, 2009. Esqueleto parcial. La mandíbula muestra un mentón con un cuerpo mandibular grácil. El poscráneo parece moderno.
- Frente a la creencia habitual de unas rutas de migación costeras, para Feng Li et al (2019), es posible que las migraciones cruzasen el centro y el norte de Asia.
Se han postulado tres modelos para explicar el desarrollo de las poblaciones del este de Asia:
- Las poblaciones del norte de Asia emigraron hacia el sur, y se mezclaron con los antepasados que se habían establecido en el Sudeste de Asia.
- El segundo modelo sugiere que las poblaciones del norte de Asia Oriental evolucionaron a partir de los pobladores del sur.
- El tercer modelo supone que las poblaciones del norte y el sur de Asia oriental evolucionaron independientemente desde finales del Pleistoceno Superior. Los dientes de Daoxian (Wu Liu et al, 2015) son muy derivados y por tanto diferentes a los hallados más al norte lo que puede apoyar la presencia de dos demos bien diferenciadas. La inestabilidad climática y la la extensión fluctuante de los desiertos (Boivin et al, 2013; Dennell, 2013) podrían haber favorecido un patrón de fragmentación y aislamiento dentro de China, dando como resultado la persistencia de linajes de Homo más primitivos en latitudes septentrionales mientras que linajes más derivados llegaron y/o evolucionaron en el sur (María Martinón-Torres et al, 2017).
- Según Dennell et al (2020) el HAM llega al sur de China desde el sudeste asiático hace ca 80 ka y al norte hace 45 ka procedente de Mongolia y el sur de Siberia, como parte de una fauna paleártica adaptada al frío. Estas poblaciones eran en gran medida independientes entre sí, y cada una de ellas debe verse como parte de sus respectivos reinos biogeográficos. La historia demográfica posterior de China es una de mezcla e interrelación de poblaciones tanto del norte como del sur de China. En el LGM, el norte de China (y Mongolia) se despoblaron en gran medida, y la posterior recolonización del norte de China se produjo tanto desde el norte como desde el sur.
En cuanto al Norte de Asia, Pitulko et al (2016) han hallado una carcasa de mamut con marcas antrópicas pre y post mortem en el paralelo 72ºN, en el río Yenisei, así como restos de un lobo cazado datados hace 45 ka, pero no se puede precisar a qué población pertenecían aquellos humanos. Disponemos de hallazgos arqueológicos datados en hace 45-40 ka, representados por sitios de las estribaciones del norte de las montañas Saian y la meseta Lena-Angara en las partes altas de los ríos Enisei, Angara y Lena. Además de los restos arqueológicos, un único fémur de Homo sapiens, que data de hace 45 ka, se encontró en Ust 'Ishim en Siberia occidental (Fu et al, 2014). De acuerdo con los hallazgos genéticos, los antepasados de la población inicial de Siberia se separaron de los primeros asiáticos orientales antes de llegar a Siberia (Fu et al, 2014; Raghavan et al, 2014).
No hay comentarios:
Publicar un comentario