 |
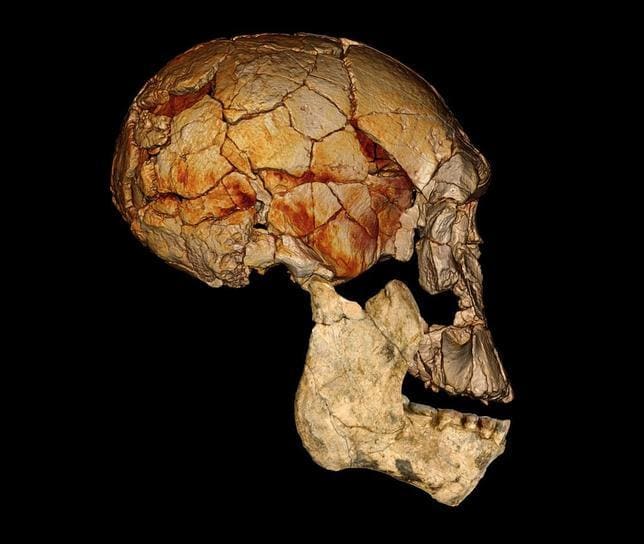
| ER 1470 combinado con ER 60000 |
Homo rudolfensis, Alexeev, 1986.
Hallazgos en Kenia (Koobi Fora), Malawi, y Etiopía (Hadar).
En 1972 Bernard Ngeneo, del equipo de Richard Leakey halló ER 1470, con un cerebro mayor, esqueleto facial más grande, ancho y plano, torus poco marcado y mandíbula y dientes mayores con grandes muelas de esmalte grueso, similares a las de los parántropos. Junto a él, ER 1472 (un fémur derecho) ER 1475 (fragmento proximal de fémur derecho), un fémur izquierdo, fragmentos distales y proximales de una tibia izquierda y ER 1481, una fíbula distal izquierda. Leakey los atribuyó a Homo sp. indet. (Leakey, 1973). Posteriormente se atribuyó a Homo habilis.
En 1972 Bernard Ngeneo, del equipo de Richard Leakey halló ER 1470, con un cerebro mayor, esqueleto facial más grande, ancho y plano, torus poco marcado y mandíbula y dientes mayores con grandes muelas de esmalte grueso, similares a las de los parántropos. Junto a él, ER 1472 (un fémur derecho) ER 1475 (fragmento proximal de fémur derecho), un fémur izquierdo, fragmentos distales y proximales de una tibia izquierda y ER 1481, una fíbula distal izquierda. Leakey los atribuyó a Homo sp. indet. (Leakey, 1973). Posteriormente se atribuyó a Homo habilis.
Para algunos, constituye un nueva especie (Homo rudolfensis o Kenyanthropus rudolfensis).
Para Alexeev (1967) y Wood (1993) los individuos de Olduvai serían habilis mientras el resto serían de otra
especie que Alexeev llamó Homo rudolfensis (de Lago Rodolfo, nombre colonial del Lago Turkana, en cuya cuenca se encontraron la mayor parte de los fósiles). Rudolfensis es más grande que habilis.
Rightmire (1995) no acepta rudolfensis, pero defiende la presencia de una especie más pequeña Homo sp., tanto en Olduvai como en Koobi Fora. Una revisión de 2007 situó los hallazgos atribuidos a rudolfensis dentro del rango del habilis.
En 2012 Meave Leakey et al, relacionaron tres fósiles hallados recientemente (ER 60000, ER 62000 y ER 62003) con ER 1470. Estos hallazgos suponen una confirmación de la especie.
 |
| Situación de Koobi Fora |
Josephine C.A. Joordens et al (2013) han datado las áreas 105 y 131 (miembro Burgi Superior) de las Cordilleras Karari y Koobi Fora (cuenca este de Turkana, Kenia) por magnetoestratigrafía y estratigrafía isotópica de estroncio, identificando la base del subcrón Olduvai además de un breve intervalo aislado de polaridad constante normal que han interpretado como un evento Pre-Olduvai.
Según los resultados, la edad de los fósiles allí hallados de Homo rudolfensis, está limitada entre 1,945 ± 0,004 y 2,058 ± 0,034 Ma. Por otra parte, hallaron una raya de origen marino en los depósitos de 2,058 ± 0,034 Ma, lo que muestra la conexión de la cuenca del Turkana con el Océano Indico, posibilitando la migración de la fauna entre estas áreas y apoyando el origen de Homo en la franja costera.
Según los resultados, la edad de los fósiles allí hallados de Homo rudolfensis, está limitada entre 1,945 ± 0,004 y 2,058 ± 0,034 Ma. Por otra parte, hallaron una raya de origen marino en los depósitos de 2,058 ± 0,034 Ma, lo que muestra la conexión de la cuenca del Turkana con el Océano Indico, posibilitando la migración de la fauna entre estas áreas y apoyando el origen de Homo en la franja costera.
 |
| Datación de las áreas 105 y 131 de Koobi Fora |
 |
| Litoestratigrafía del Área 131 de Koobi Fora |
Holotipo
KNM-ER 1470. 1972. Koobi Fora, Kenya. 1,9 Ma. Bernard Ngeneo, del equipo de Richard Leakey. Se trata del cráneo de rudolfensis más completo por ahora. Su capacidad craneal es de 750 cc, grande para un habilis. En un principio se le estimó una edad de 3 Ma y se confundió con un australopiteco. Su bóveda craneal es más grande y menos robusta que la de los australopitecos. Carece de cresta craneal. Su cara es prominente y robusta. Una reconstrucción por ordenador realizada en 2007 determinó que el cráneo no se había reconstruido de forma adecuada y redujo su volumen a 575 cc, por lo que no tendría nada de especial y estaría dentro del rango de habilis.
Cráneos
- KNM-ER 1470.
- KNM-ER 1590. Wood, 1991. Incluye una parte importante de la calvaria, que se parece mucho a la de 1470 en tamaño y forma, y restos dentales de gran tamaño que, por ser dientes diferentes o en distinto grado de desarrollo, no pueden ser comparados con los de 1470.
- KNM-ER 3732. Wood, 1991. Cráneo parcial, robusto, con la parte superior de la cara y cigomático izquierdo. Los huesos frontal y parietal proporcionan una indicación general de mayor capacidad. En las regiones supraorbital y orbital difiere de 1470.
- KNM-ER 3891. Wood, 1991. Fragmentos craneales incluyendo fragmentos de maxilar.
- KNM-ER 62000. Cráneo bien preservado de un joven de unos ocho años, con algunos dientes. Cara larga y aplanada. Datado entre 1,95-1,78 Ma.
Mandíbulas
Los restos que a continuación se detallan son consistentes con KNM-ER 1470, pero no puede establecerse una relación directa. En algunas características, son similares a Paranthropus boisei.
- KNM-ER 819. Wood, 1991. Fragmento de mandíbula.
- KNM-ER 1482. Wood, 1991. Mandíbula.
- KNM-ER 1483. Wood, 1991. Fragmento de mandíbula.
- KNM-ER 1801. Wood, 1991. Fragmento de mandíbula.
- KNM-ER 1802. Wood, 1991. Está razonablemente completa. Muestra marcado relieve en el cuerpo externo y eversión de la base.
- KNM-ER 60000 Mandíbula inferior casi completa de un adulto, con varios dientes y raíces. Datada entre 1,95-1,78 Ma.
- KNM-ER 62003 Fragmento de mandíbula inferior, con varios incisivos. Datado entre 1,95-1,78 Ma.
 |
| UR 501 |
- UR 501. Chiwondo Beds, Uraha, Malawi, ca. 2.5-1.9 Ma?. Mandíbula relativamente completa con sus dientes, de edad incierta. Premolares más pequeños que KNM-ER 1802. La inclusión en Homo se basa en la morfología molar y premolar y en la anatomía mandibular. Ha sido relacionada con KNM-ER 1802 (Homo rudolfensis; Bromage et al, 1995) basándose en el gran tamaño de los dientes anteriores y poscaninos, el esmalte molar relativamente grueso, y las raíces del P4 dobles, planas y bajas.
Restos poscranelaes
Relacionados con KNM-ER 1470 solo en base a su tamaño.
- KNM-ER 813. Astrágalo.
- KNM-ER 1472 (Wood, 1992). Fémur.
- KNM-ER 1473. Húmero proximal.
- KNM-ER 1481 A/B (Wood, 1992). Fémur izquierdo completo, ambas terminaciones de la tibia izquierda y la terminación inferior del peroné izquierdo, similares a los de los humanos modernos.
- KNM-ER 3228 (McHenry y Coffing, 2000). Coxis.
PRINCIPALES FÓSILES
|
|||
YACIMIENTOS
|
CRÁNEOS
|
MANDÍBULAS
|
POSCRÁNEOS
|
Koobi
Fora, Lago Turkana, Kenia
|
ER 1470
ER 1590
ER 1813
ER 3732
ER 3891
ER 62000
|
ER 819
ER 1482 ER 1483
ER 1501
ER 1801
ER 1802
ER 1805
ER 1813
ER 60000
ER
62003
|
ER
813
ER
1472
ER
1481
|
Malawi
|
UR 501
|
||
No está muy clara la clasificación de la mandíbula
de Malawi (2,4 Ma) y dientes aislados de Omo.
Morfología
Las características morfológicas se parecen en unos aspectos a habilis y en otros a los parántropos (Wood, 2000).
- Cerebros grandes. Capacidad media 750 cc. El cráneo de KNM-ER 1470 muestra un ligero torus supraorbital sin sulcus y una constricción retroorbital moderada. La fosa glenoide y el meato acústico externo están en una posición más avanzada que en los Australopithecus y se observa una cara larga con la parte superior estrecha. El maxilar es cuadrado, con un paladar plano y muy corto que presenta una gran anchura que contrasta con la de los australopitecos. Los rebordes del orificio piriforme son redondeados y están inclinados. El arranque del arco cigomático se inclina hacia delante.
- Dentición poscanina bastante grande, con un cociente de megadoncia de 1,5, similar a erectus. Grandes coronas y raíces más complejas que en habilis, con el último molar más grande que el segundo. Grandes alveolos sugieren potentes caninos e incisivos.
- Los escasísimos materiales poscraneales sugieren una bipedación más lograda que la de habilis (Picq, 2004).
 |
| Diferencias principales entre rudolfensis y habilis. Niguna de estas especies puede ser la antecesora de los últimos Homo, ya que no presentan simultáneamente rasgos derivados en el cráneo, aparato masticatorio y postcráneo. |
Homo rudolfensis
|
Homo habilis
| |
Espesor de la bóveda craneal
|
espesa
|
delgada
|
Depresión trasera en la región orbital
|
ausente
|
presente
|
Depresión postorbital lateral
|
ausente
|
presente
|
Posición de las líneas curvas en los temporales
|
mediana
|
alta
|
Forma de la porción escamosa del hueso temporal
|
alta y redonda
|
baja y triangular
|
Orientación del borde anterior de la porción escamosa del hueso temporal
|
anterior
|
vertical
|
Unión de crestas mastoideas y supramastoideas
|
presencia de unión
|
ausencia de unión
|
Inclinación de la región nucal
|
débil
|
fuerte
|
Torus occipital
|
ausente
|
desarrollo medial
|
Forma de la raíz del proceso cigomático del hueso temporal
|
plana
|
elíptica
|
Anchura de la cara superior / cara media
|
más ancha
|
idéntica
|
Posición del punto frontomaxilar respecto a la fila de dientes
|
P4-M1
|
M1
|
Tamaño del hueso cigomático
|
grande
|
corto
|
Fosa cigomático-maxilar
|
ausente
|
presente
|
Individualización de la región de los incisivos con respecto a la abertura nasal
|
la región de los incisivos participa en la abertura nasal
|
la región de los incisivos es independiente
|
Forma de las órbitas
|
cuadrada
|
rectangular con alargamiento supero-inferior
|
Posición del borde superior de la órbita con respecto al borde inferior
|
en el mismo plano
|
en posición anterior
|
Superficie oclusal del tercer molar / segundo molar
|
>1
|
<1
|
Tamaño de los incisivos
|
grande
|
pequeño
|
Forma de la región mentoniana
|
subrectangular
|
convexidad medio-lateral
|
Paralelismo del eje largo de la mandíbula y de la fila de dientes
|
paralelos
|
< 15°
|
Discusión filogenética
- Wood y Collar (1990) han propuesto incluir Homo habilis y Homo rudolfensis en el género Australopithecus: El esqueleto postcraneal de OH 62 es prácticamente idéntico al del Australopithecus afarensis, con conservación de la conducta trepadora. Podría incluirse también Kenyanthropus platyops (Cela y Ayala, 2003, consideran que deben clasificarse juntos).
- Cela y Ayala (2006) entienden que sería convertir Australopithecus en un cajón de sastre imposible de ordenar.
- Otra alternativa es incluir estos taxones en el género Kenyanthropus.
- La tercera, recomendada por Cela y Ayala, es incluirlos en el género Homo. Platyops sería la species germinalis.
- El parecido con Paranthropus robustus en el aparato masticatorio y la dentición hace que algunos paleoantropólogos hayan vinculado a rudolfensis con los parántropos. Otros consideran que estos parecidos son homoplasias.
Otros hallazgos de cronología similar pero difícil clasificación
- KNM-ER 1805, The Mystery Skull. 1973. Koobi Fora, Kenya. 1,85 Ma. Paul Abell. Buena parte de un cráneo incluyendo muchos dientes. 600 cc. Con una cresta sagital parecida a la de P. boisei, pero con los dientes demasiado pequeños para pertenecer a esta especie.
- Podría ser asignado a casi cualquier especie pero parece más similar a Homo habilis (Wood, 1991).
- Un reciente estudio cladístico, lo colocó dentro de los australopitecinos robustos si bien diferente a cualquier especie descrita.
 |
| Reconstrucciones virtuales de KNM-ER 1805 (A) y OH 24 (B). En B las partes grises representan elmentos completados por espejo. Escala 5 cm. David Lordkipanidze et al, 2013. |
- KNM-ER 1813. 1973. Koobi Fora. 1,9-1,8 Ma. Kamoya Kimeu. Similar al 1470, pero mucho más pequeño. 510 cc, muy pequeño para habilis. Algunos creen que es un caso de dimorfismo sexual otros creen que es una especie diferente de Homo y otros que es un australopitecino. Sorprendentemente moderno, sin cresta sagital y con prominencia nasal pequeña.
- Ferguson (1995) lo convirtió en el holotipo de Homo microcranous. Candidatos a paratipos serían KNM-ER 1501, 1805, 3735, 42703; OH 13, 24, 62 y 65.
- Para Howell (1998) y Wood (1991) pertenece a Homo habilis.
 |
| Reconstrucción digital de KNM-ER 1813. Stefano Benazzi et al (2013) |
- KNM-ER 3735. 1975. Área 116, Koobi Fora. 1,9-1,88 Ma. Kamoya Kimeu. Esqueleto de adulto, posiblemente macho: 14 fragmentos craneales, clavícula, escápula, radio, ulna, dos falanges de la mano, sacro, fémur y tibia. Brazos muy grandes en relación al cuerpo y potente cintura escapular, lo que parece implicar la retención de rasgos simiescos (Haeusler y McHenry, 2007).
- Tobias (1989), lo incluyó en Homo habilis.
- Para Bernard Wood (1991) se trata de Homo sp. indet.
- KNM-ER 5881. En 1980, Koobi Fora proporcionó un fragmento de fémur de Homo datado en 1,88 Ma. En 2009 se recuperaron otro fragmento y un fragmento de ilion del mismo individuo.
- KNM-ER 42703. 2000. Área O, Ileret. 1,44 Ma. John E. Kaatho. Maxilar derecho que incluye parte del palatino, proceso cigomático, la mayor parte del proceso alveolar y dientes de C a M3 superior. Con unos molares más pequeños y un M3 superior mas corto mesodistalmente, se separa claramente de habilis y erectus.
- William L. Jungers et al han comunicado en la 84a Reunión Anual de la Asociación Americana de Antropólogos Físicos (2015) el hallazgo de nuevos fósiles de homínidos en Kolom Odiet, Ileret, cuenca del Turkana, Kenia, durante las expediciones de campo de 2012-2013:
- KNM-ER 64060.
- Una dentición inferior con todos los dientes salvo el incisivo central derecho. 2,3-2,2 Ma.
- Se atribuye al género Homo, sobre la base de la morfología y la métrica. Posiblemente, Homo habilis.
- Presenta unos caninos de los más grandes registrados en los primeros Homo.
- KNM-ER 64061.
- Esqueleto parcial. 2,3-2,2 Ma. Conserva la mayor parte de ambas diáfisis humerales, un cúbito derecho casi completo, gran parte de la clavícula derecha y la cavidad glenoidea de la escápula derecha.
- Los huesos largos son relativamente gráciles pero poseen corticales gruesos en sección transversal.
- En general, la morfología sugiere que su pertenencia a un Homo temprano. Posiblemente erectus.
- KNM-ER 64062.
- Esqueleto parcial. 1,86-1,82 Ma. Conserva un húmero distal, un escafoides izquierdo y un pie derecho parcial que incluye un calcáneo, astrágalo, cuboides, navicular, cuneiformes intermedios y laterales, el metatarso hallucal y su falange proximal, y partes de los cuatro metatarsianos menores.
- El escafoides se asemeja al de otros homínidos tempranos (por ejemplo, STW 618, LB 1). Los huesos de los pies muestran una mezcla de características primitivas y derivadas. El hallux y el astrágalo son similares a los de Dmanisi; el escafoides luce una gran tuberosidad y está aplastado lateralmente. El calcáneo tiene un tubérculo inflado con ambos procesos plantares y al igual que el cuboide tiene un aspecto moderno.
- En general, la morfología sugiere su pertenencia a un Homo temprano. Posiblemente erectus.
Adam van Arsdale
Bibliografía:
- Leakey, Richard E. Koobi Fora research project. Clarendon Press, 1978.
- Walker, Alan, and RICHARD E. Leakey. "The hominids of East Turkana." Scientific American 239.2 (1978): 54.
- Feibel, Craig S., Francis H. Brown, and Ian McDougall. "Stratigraphic context of fossil hominids from the Omo Group deposits: Northern Turkana Basin, Kenya and Ethiopia." American Journal of Physical Anthropology 78.4 (1989): 595-622.
- Wood, B. A. "Koobi Fora Research Project IV: Hominid Cranial Remains from Koobi Fora." Oxford: Oxford University Press.——(1992)‘Origin and evolution of the genus Homo’, Nature 355 (1991): 783-90.
- Schrenk, F., et al. "Early hominid diversity, age and biogeography of the Malawi-Rift". Human Evolution 17.1-2 (2002): 113-122.
- Feibel, Craig S., Christopher J. Lepre, and Rhonda L. Quinn. "Stratigraphy, correlation, and age estimates for fossils from Area 123, Koobi Fora." Journal of human evolution 57.2 (2009): 112-122.
Te puede interesar:
|
Dating for everyone is here: ❤❤❤ Link 1 ❤❤❤
ResponderEliminarDirect sexchat: ❤❤❤ Link 2 ❤❤❤
il. .