Se han encontrado abundantes fósiles de hominoideos datados en el Mioceno Medio (16-10 Ma) en Asia y Europa, pero pocos en África. Sin duda, la colonización del suelo del bosque constituyó un éxito para los hominoideos que, desde África, se extendieron por Eurasia donde las condiciones ambientales les eran más favorables.
- Entre 14-12 Ma la temperatura del planeta descendió unos 12ºC de media. Los casquetes polares se hicieron muchos más grandes y consiguientemente bajó el nivel del mar. En África las áreas boscosas dejaron paso a espacios abiertos y secos.
- En Eurasia, el clima comienza a estar marcado por las estaciones. Disponiendo de mayor humedad que en África, los bosques se extienden por el Sur de Eurasia. Los hominoideos engrosan el esmalte dentario y robustecen las mandíbulas, colonizan el suelo de bosques y selvas y consumen semillas, tubérculos y raíces, además de frutos, hojas y yemas.
- Las condiciones climáticas y ecológicas permiten un cambio a a dietas de frutas maduras ricas en fructosa pero con bajo contenido de vitamina C, con pérdida de la encima uricasa. La capacidad de aumentar el volumen de grasa cuando la fruta madura es abundante proporcionaría un amortiguador para la supervivencia durante la escasez.
- En este ambiente, se fueron separando los linajes, cada vez más especializados, engrosando el esmalte dentario y robusteciendo las mandíbulas. Cuando una especie encuentra nichos ecológicos vacíos, puede especializarse para llenarlos. Este fenómeno se denomina radiación adaptativa.
 |
Hominoideos de Eurasia occidental. Isaac Casanovas-Vilar et al. 2011.
|
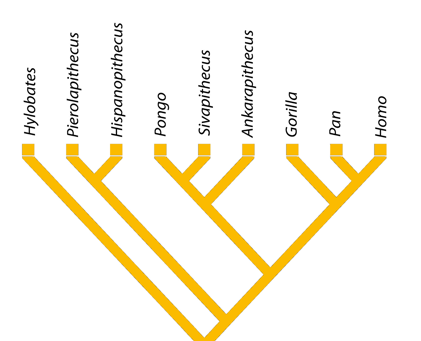 |
| Árbol filogenético de la rama de los hominoideos a la que pertenecemos los humanos. Miriam Pérez de los Ríos. |
En el árbol filogenético de los hominoideos, se conservan
durante el Mioceno Medio las ramas correspondientes a los Proconsulidae,
Afropithecidae Hylobatidae y Hominidae. Esta última rama surgida en el Mioceno Inferior se
divide, a su vez, en varias subfamilias:
- Homininae: La rama que dará origen a gorilas, chimpancés y seres humanos, con especies totalmente adaptadas al suelo, aunque conservan capacidad de trepa. No se han hallado fósiles correspondientes a este periodo.
- Ponginae. La que dará lugar a los orangutanes (póngidos), con fósiles hallados principalmente en Asia. Según evidencias genéticas, la separación se produjo hace 14 Ma. Aunque prefieren los árboles, los orangutanes pueden caminar por el suelo apoyando las falanges proximales de las extremidades anteriores (fist walking).
- Sivapithecus
- La morfología craneal es similar a la de Pongo. Cráneo con órbitas altas y redondeadas, separadas entre sí por un tabique muy estrecho. La frente se extendía verticalmente, sin reborde óseo, como en los orangutanes. Esmalte grueso, lo que sugiere una dieta de nueces, semillas y otros vegetales duros. Cuadrúpedo. Dimorfismo sexual apreciable en la talla y en los caninos.
- La morfologío postcraneal de Sivapithecus difiere en muchos detalles de la de las especies existentes. No hay características postcraneanas sinapomórficas obvias compartidas con los hominoideos existentes, incluyendo Pongo. La morfología corresponde a un movimiento lento, pausado y arbóreo por encima de soportes combinado con un comportamiento antipronógrado frecuente.
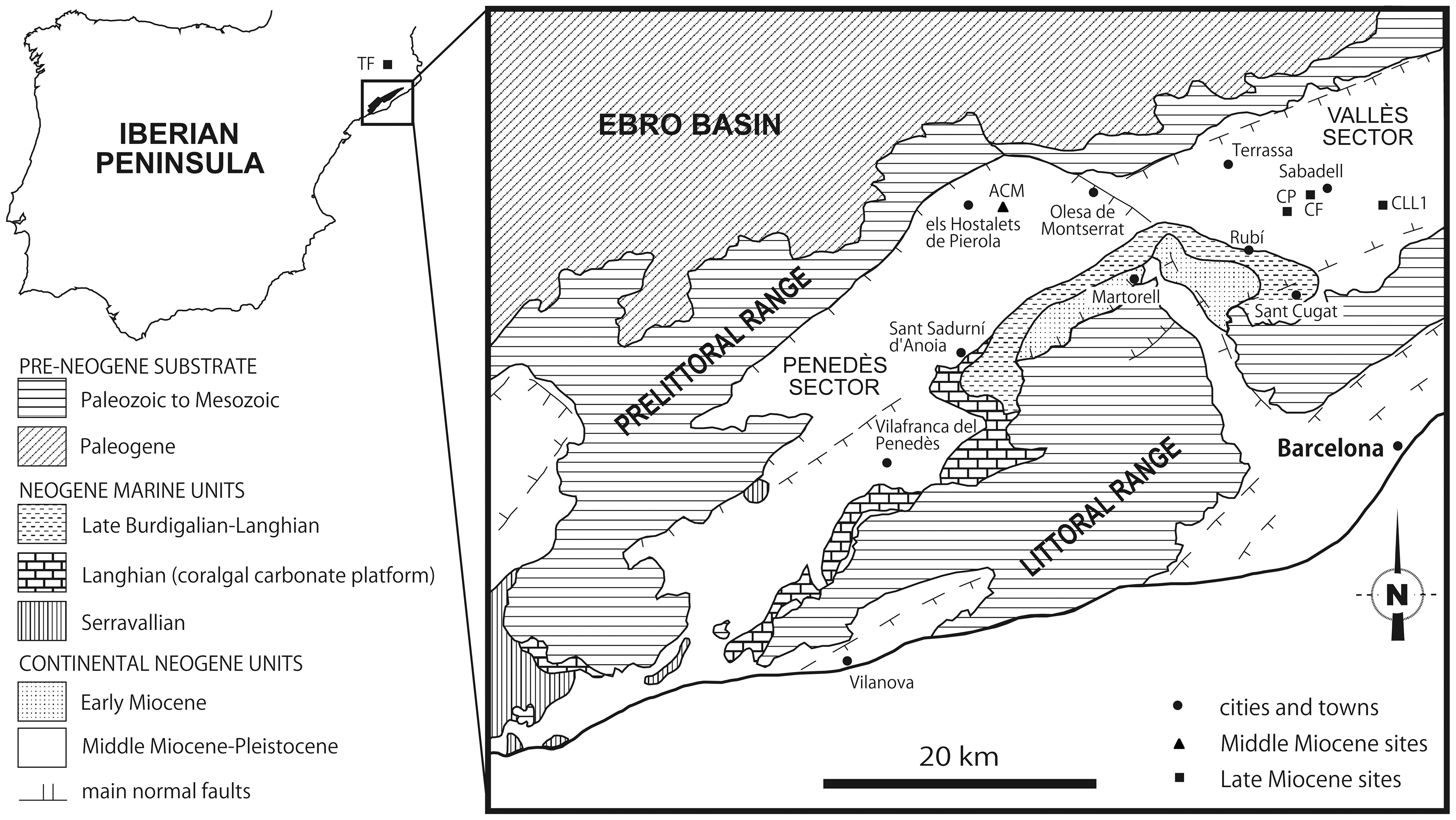 |
| Yacimientos de hominoideos en los alrededores de Barcelona. Daniel DeMiguel, David M. Alba y Salvador Moyà-Solà, 2014. |
- Kenyapithecinae.
- En 1999 Ward y colaboradores describieron un esqueleto parcial hallado en Kipsaramon (Muruyur, Tugen Hills, Baringo, Kenya) identificado como KNM-TH 28860. Es el más antiguo ejemplar de un hominoideo en el que se localizan rasgos avanzados tanto en el aparato masticatorio como en el locomotor. Fue datado en algo más de 15 Ma. Constituye el holotipo de Kenyapithecus wickeri. Dividía su tiempo entre el suelo y los árboles.
- Este hallazgo refutó la hipótesis de Andrews según la cual existieron dos líneas evolutivas separadas, una de ellas con rasgos evolucionados en el aparato masticatorio y arcaicos en el locomotor y otra al contrario.
- El pie de Kenyapithecus africanus (KNM-MB 31335; Maboko Island) presenta la morfología de plantígrado más temprana. El talón toca primero el suelo durante la locomoción terrestre, un rasgo que solo comparten los simios africanos y los humanos. KNM-MB 28397 presenta otra característica avanzada: el alargamiento, orientación plantar y extensividad lateral del tubérculo peroneal del quinto metatarsiano, el cual está orientado plantarmente. Datados en 15 Ma. Irisa D. Arney et al (2013).
- Dryopithecinae. Con características intermedias entre los póngidos (Sivapithecus) y los kenyapithecinos.
- En 2003 el equipo de Salvador Moyà-Solà encontró en Can Vila, Hostalets de Pierola, Barcelona, 83 huesos o fragmentos de un primate de 35 kg y alimentación a base de frutas: IPS 21350, holotipo de Pierolapithecus catalaunicus. Cara ancha y plana, largo morro con grandes caninos, dedos cortos que indican buena aptitud para trepar y correr por los árboles. Caja torácica ancha y amplia, con los omóplatos a la espalda y no a los lados. Columna vertebral más corta y rígida, especialmente en la zona lumbar. Brazos largos. Desplazamiento colgándose de las ramas (postura ortógrada); primera evidencia conocida, aunque también presenta adaptaciones primitivas para el desplazamiento cuadrúpedo por encima de las ramas. De los restos de pelvis (fragmento de ileon y pequeño fragmento del acetábulo caudal del isqueon; los más antiguos conservados de un simio) se deduce que podía caminar erguido y trepar verticalmente. Estos rasgos relativos a la forma de desplazamiento debieron evolucionar en paralelo con los que presenta la rama que dará lugar a los humanos (homoplasia); se descarta la herencia de un ancestro común. Datado en 11,9 Ma. (Sergio Almécija et al, 2009; Ashley S. Hammond et al, 2013; Marta Pina et al, 2014).
- Pierolapithecus carece de seno frontal, y sus canales lacrimales se desarrollan hacia adelante como en los póngidos y no verticalmente como en los grandes simios africanos. Otras características como la base de la cavidad nasal –que incluye el paladar- apuntan a un parecido más directo con los driopitecinos. Por otra parte, los turbinales, láminas óseas que se desarrollan en la cavidad nasal para sustentar las partes blandas de la nariz, se desarrollan de una manera muy similar a Pongo. Todos estos datos muestran que Pierolapithecus catalaunicus presentaba un mosaico de características propias de los homínidos primitivos, así como algunas derivadas de los ponginos, que son inconsistentes con la hipótesis que Pau fuera uno de los primeros homínidos (Miriam Pérez de los Ríos, 2012).
- Vivió en bosques de laurisilva (hojas grandes y lanceoladas, perennes).
- Anoiapithecus
- Anoiapithecus brevirrostris
- Lluc, rostro parcial con mandíbula.
- IPS 35027: fragmento de maxilar con M1–M2 derechos y P4–M2 izquierdos, datado en 12,3–12,2 Ma. Hallado en Abocador de Can Mata (Barcelona). Asignación taxonómica en estudio.Representa el fósil hominoideo más antiguo hallado en la Península Ibérica.
- IPS 41712: fragmento de maxilar izquierdo con P3–M1, datado en 11,9 Ma.
- Dryopithecus pesaba entre 20-30 kg, con un acusado dimorfismo sexual. Caninos desarrollados en los machos. Molares de esmalte delgado, lo que indica alimentación de frutos carnosos. Brazos más largos que las piernas y primeras y segundas falanges de los dedos de las manos extraordinariamente largas, lo que indica locomoción suspendida, similar a la de los orangutanes. Yacimientos en Vallès-Penedès, Udabno (Cáucaso) y Rudabanya (Europa Central).
- Dryopithecus fontani. 11.9-10.5 Ma.
- Abocador de Can Mata en els Hostalets de Pierola. Cara parcial de un individuo adulto de sexo masculino (ACM/C3-Ae; 11,9 Ma). La morfología en algunos aspectos recuerda la de los gorilas actuales.Un canino superior de macho (ACM/C4-Ap; 11,9 Ma). Un fragmento proximal de fémur´(ACM/C3-Az; 11.9 Ma, atribuido provisionalmente a D. fontani en base a su gran tamaño.
- Castell de Barberà en Barberà del Vallès (ca. 11.2-10.5 Ma) dos fragmentos de falanges pollicales y una diáfisis humeral parcial. Asignación dudosa.
- La Grive-Saint-Alban.
- Oreopithecus habitaba la isla tosco-sarda. Entre 20-30 kg. Cráneo pequeño y globuloso. Mandíbula robusta. Maxilar corto con caninos de pequeño tamaño. Premolares con dos cúspides. Esmalte delgado, con gran número de crestas y pequeñas cúspides, lo que indica una dieta basada en hojas. Pies prensiles y brazos mucho más largos que las piernas, lo que indica capacidad de trepa y suspensión, similar a la de los orangutanes. Gabrielle A. Russo y Liza J. Shapiro (2013) han reinterpretado los restos postcraneales BA # 72 (Oreopithecus bambolii) procedentes de Baccinello, Toscana. Un estudio anterior, había encontrado evidencias de bipedación, a partir de la pelvis y la curvatura lumbar (Köhler y Moyà-Solà, 1997), pero según el neuvo estudio, la región lumbosacra no presenta adaptaciones para la bipedación. Según Sergio Amécija et al (2014) poseía mayor capacidad de manipulación que los simios no humanos existentes. Frank L’Engle Williams (2013), a partir del microdesgaste dental del ejemplar IGF 11778, propone una dieta folívora con forrajeo intermitente extractivo y/o terrestre. A partir de los isótopos de carbono y oxígeno en el esmalte de los dientes, para Nelson y Rook (2016) Oreopithecus habitaba bosques abiertos y su alimentación incluía probablemente tubérculos y vegetación acuática. Los valores de presencia de los isótopos son similares a Ardipithecus.
 |
| BA # 72 (Oreopithecus bambolii) |
- Hispanopithecus (Alba et al, 2018).
- Hispanopithecus laietanus (11,1-9,5 Ma) presenta características faciales y del resto del esqueleto más derivadas que los driopitecinos del Mioceno medio (Pierolapithecus, Dryopithecus y Anoiapithecus). Se trata del primer homínoideo que presenta adaptaciones para desplazarse suspendido bajo las ramas (plan corporal ortrógado), como ha mostrado el estudio del espécimen IPS18800 Jordi, a pesar de conservar aún algunas adaptaciones más primitivas para el desplazamiento cuadrúpedo arbóreo (Melissa Tallman et al, 2013).
- Fue definido originalmente sobre la base de una mandíbula inferior y restos dentales de La Tarumba 1.
- Restos dentales en varias localidades, aunque la muestra más grande proviene de Can Llobateres 1. Además, en este yacimiento, algunos huesos aislados.
- Cráneo parcial de Can Llobateres 2. Esqueleto parcial de alrededor de 60 elementos.
- Restos inéditos de la Estación Depuradora de Aguas Residuales del Ripoll, en Sabadell (EDAR).
- Esqueleto parcial de Can Feu, Sant Quirze del Vallès.
- Hispanopithecus crusafonti ca 10,4-10 Ma). Especie muy discutida.
- La mayor parte de los restos, provienen de Can Poncic 1.
- Mandíbula parcial de Teuleria del Firal.
- Hispanopithecus hungaricus (ca 10-9,8 Ma).
- Según Daniel DeMiguel, David M. Alba y Salvador Moyà-Solà (2014) la dieta de Pierolapithecus se asemejaba a los consumidores de alimentos duros, Oreopithecus forrajeaba frutas blandas y otros alimentos de forma oportunista y los demás géneros carecen de análogos dietéticos existentes claros: muestran algunos grados de esclerocarpia (abrían cáscaras duras con sus dientes), con mayor claridad en Griphopithecus y Ouranopithecus (adaptado a los ambientes más abiertos y áridos) mientras que Anoiapithecus, Dryopithecus e Hispanopithecus dependen en mayor medida de frutos blandos. Por lo tanto, en contraste con la condición esclerocárpica vigente al inicio de la radiación de los homínoideos de Eurasia, en el Mioceno Tardío la dieta de frutas blandas coexistió con la de alimentos duros. A pesar de una tendencia climática hacia el enfriamiento y el aumento de la estacionalidad, se produjo una progresiva diversificación de la dieta que no incluyó la alimentación estricta con hojas.
 |
| Pierolapithecus catalaunicus. Se destacan los restos de pelvis. |
Adam Van Arsdale. The Age of Apes.

No hay comentarios:
Publicar un comentario